

Purification of active HOPS complex reveals its affinities for phosphoinositides and the SNARE Vam7p
- PMID: 16601699
- PMCID: PMC1440844
- DOI: 10.1038/sj.emboj.7601051
Purification of active HOPS complex reveals its affinities for phosphoinositides and the SNARE Vam7p
Abstract
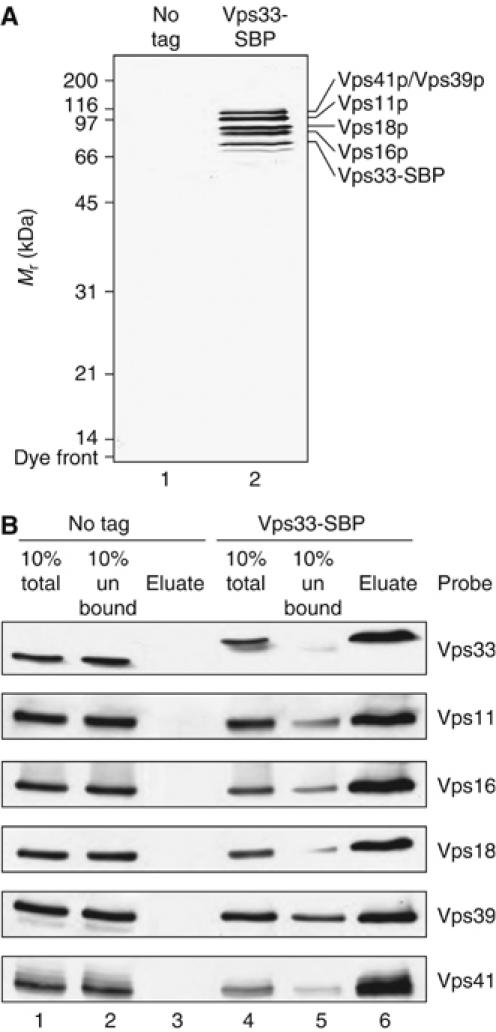
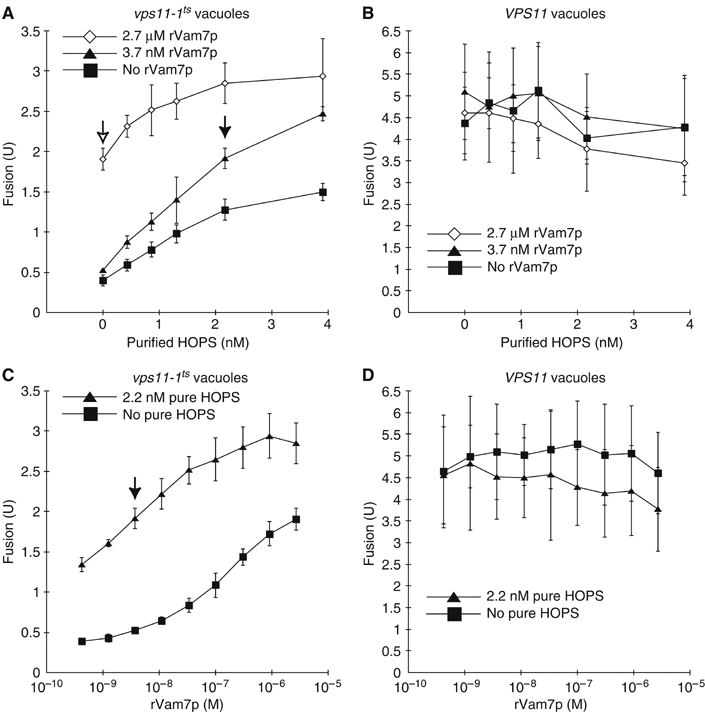
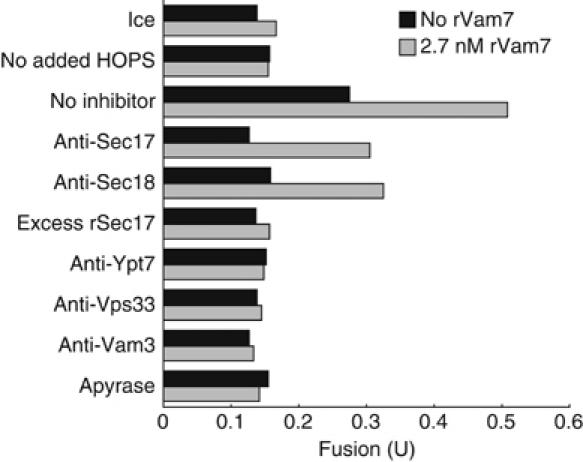
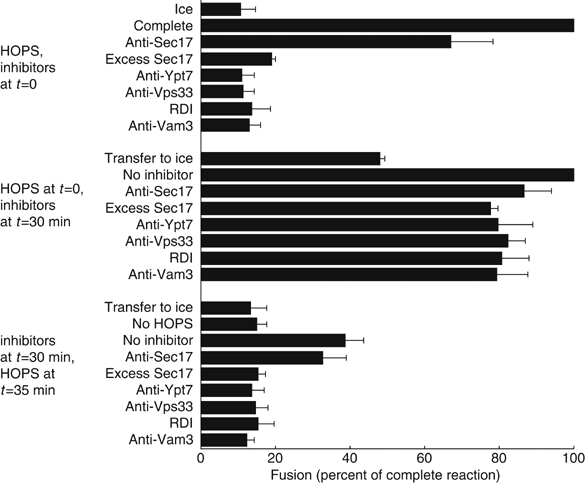
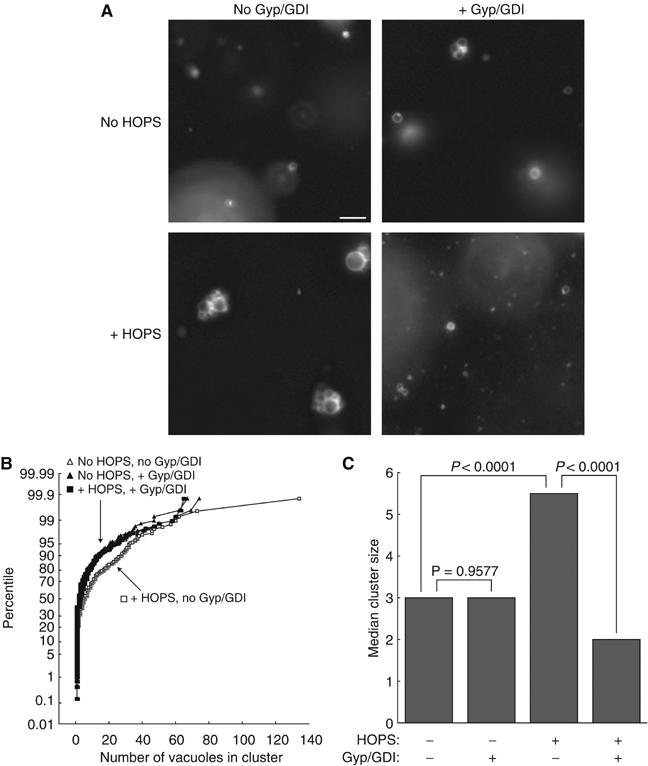
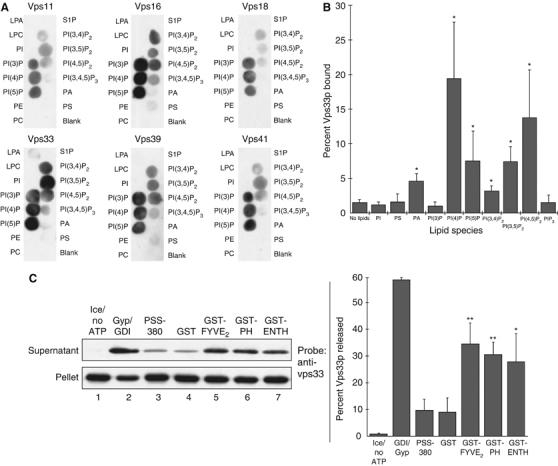
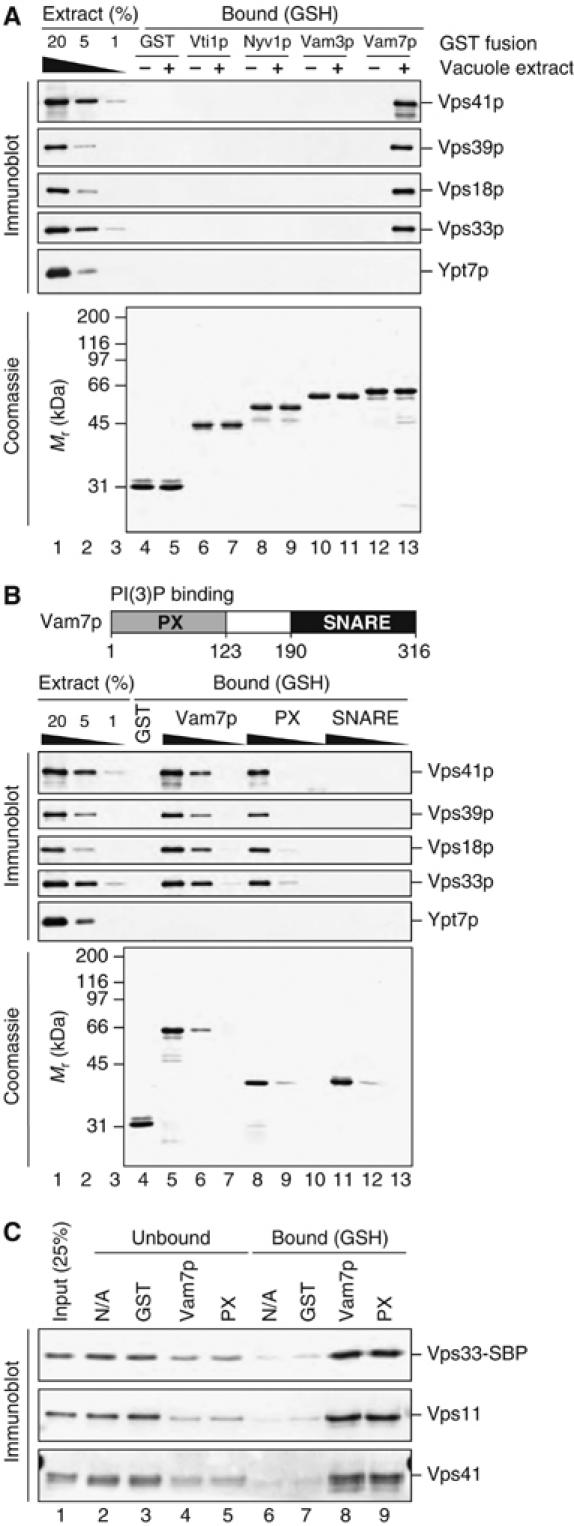
Coupling of Rab GTPase activation and SNARE complex assembly during membrane fusion is poorly understood. The homotypic fusion and vacuole protein sorting (HOPS) complex links these two processes: it is an effector for the vacuolar Rab GTPase Ypt7p and is required for vacuolar SNARE complex assembly. We now report that pure, active HOPS complex binds phosphoinositides and the PX domain of the vacuolar SNARE protein Vam7p. These binding interactions support HOPS complex association with the vacuole and explain its enrichment at the same microdomains on docked vacuoles as phosphoinositides, Ypt7p, Vam7p, and the other SNARE proteins. Concentration of the HOPS complex at these microdomains may be a key factor for coupling Rab GTPase activation to SNARE complex assembly.
Figures
References
-
- Allan BB, Weissman J, Aridor M, Moyer B, Chen CD, Yoo JS, Balch WE (2000) Stage-specific assays to study biosynthetic cargo selection and role of SNAREs in export from the endoplasmic reticulum and delivery to the Golgi. Methods 20: 411–416 - PubMed
-
- Aravind L, Iyer LM, Koonin EV (2003) Scores of RINGS but no PHDs in ubiquitin signaling. Cell Cycle 2: 123–126 - PubMed
-
- Babst M (2005) A protein's final ESCRT. Traffic 6: 2–9 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
