

Nucleolin is a histone chaperone with FACT-like activity and assists remodeling of nucleosomes
- PMID: 16601700
- PMCID: PMC1440837
- DOI: 10.1038/sj.emboj.7601046
Nucleolin is a histone chaperone with FACT-like activity and assists remodeling of nucleosomes
Abstract
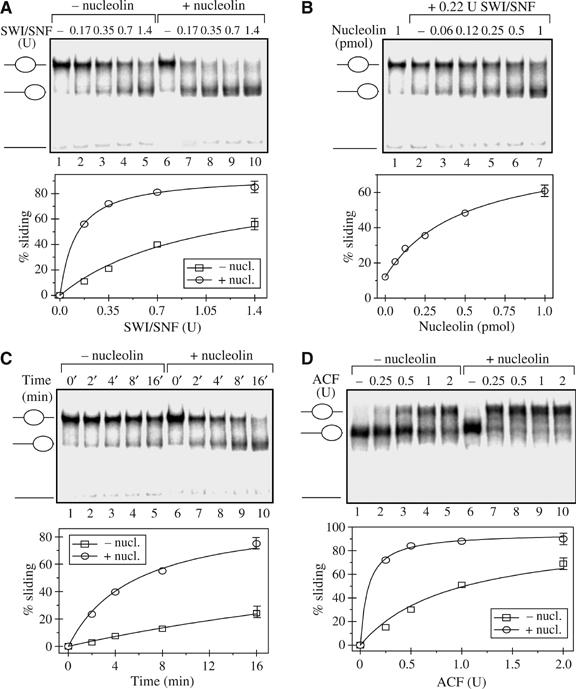
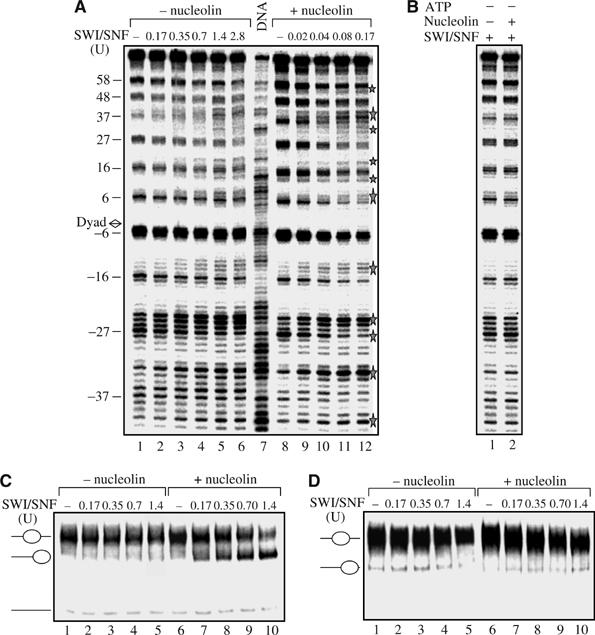
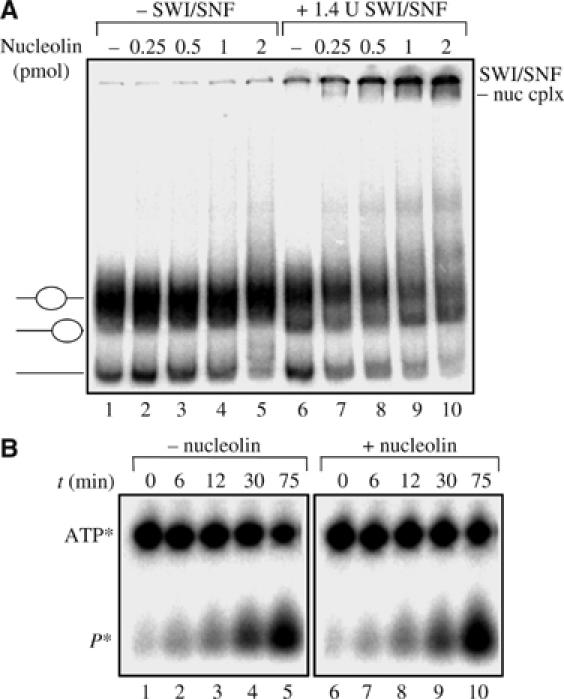
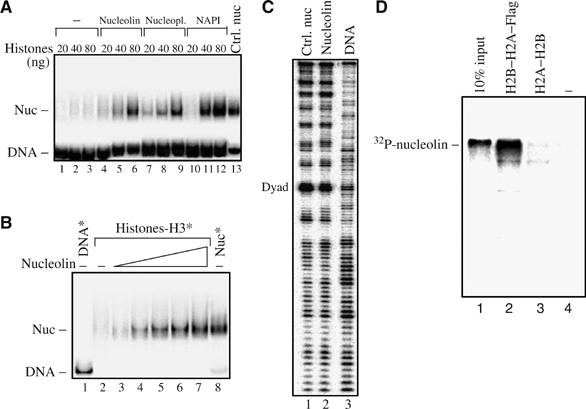
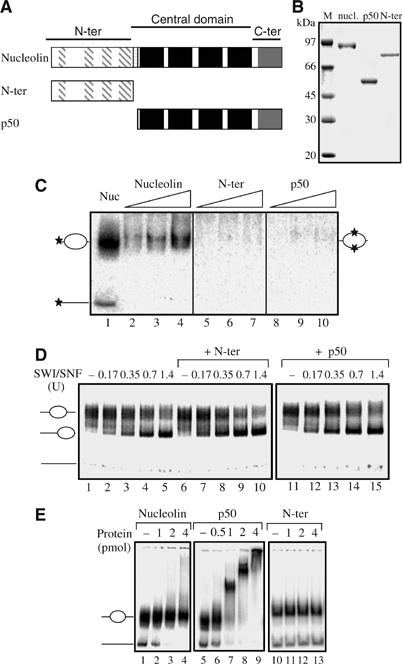
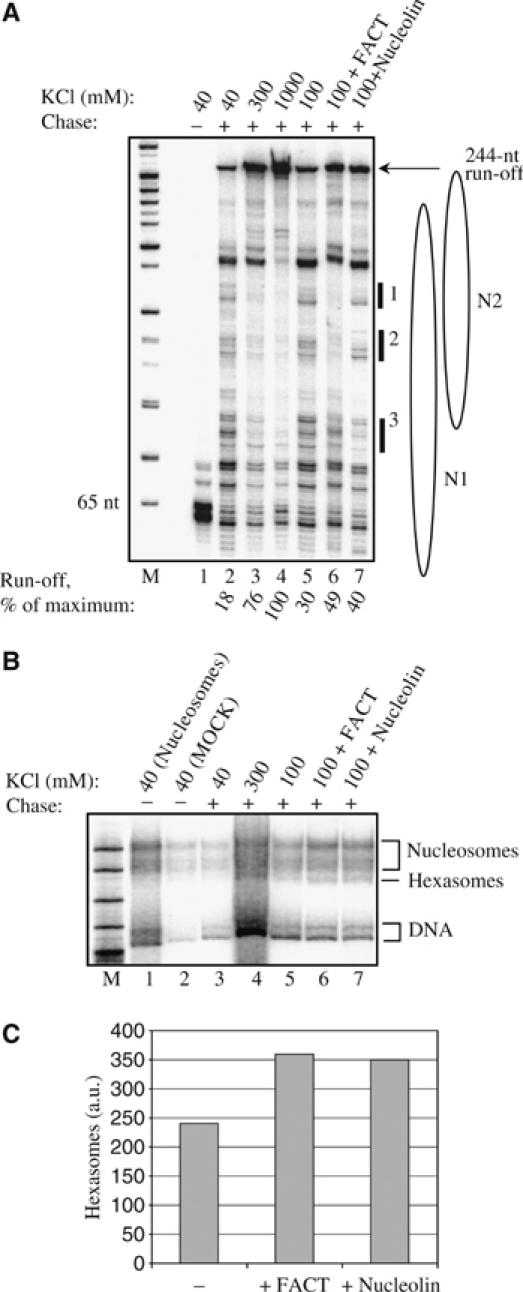
Remodeling machines play an essential role in the control of gene expression, but how their activity is regulated is not known. Here we report that the nuclear protein nucleolin possesses a histone chaperone activity and that this factor greatly enhances the activity of the chromatin remodeling machineries SWI/SNF and ACF. Interestingly, nucleolin is able to induce the remodeling by SWI/SNF of macroH2A, but not of H2ABbd nucleosomes, which are otherwise resistant to remodeling. This new histone chaperone promotes the destabilization of the histone octamer, helping the dissociation of a H2A-H2B dimer, and stimulates the SWI/SNF-mediated transfer of H2A-H2B dimers. Furthermore, nucleolin facilitates transcription through the nucleosome, which is reminiscent of the activity of the FACT complex. This work defines new functions for histone chaperones in chromatin remodeling and regulation of transcription and explains how nucleolin could act on transcription.
Figures

References
-
- Adkins MW, Howar SR, Tyler JK (2004) Chromatin disassembly mediated by the histone chaperone Asf1 is essential for transcriptional activation of the yeast PHO5 and PHO8 genes. Mol Cell 14: 657–666 - PubMed
-
- Adkins MW, Tyler JK (2006) Transcriptional activators are dispensable for transcription in the absence of spt6-mediated chromatin reassembly of promoter regions. Mol Cell 21: 405–416 - PubMed
-
- Angelov D, Lenouvel F, Hans F, Muller CW, Bouvet P, Bednar J, Moudrianakis EN, Cadet J, Dimitrov S (2004a) The histone octamer is invisible when NF-kappaB binds to the nucleosome. J Biol Chem 279: 42374–42382 - PubMed
-
- Angelov D, Molla A, Perche PY, Hans F, Cote J, Khochbin S, Bouvet P, Dimitrov S (2003) The histone variant macroH2A interferes with transcription factor binding and SWI/SNF nucleosome remodeling. Mol Cell 11: 1033–1041 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
