

Endogenous and synthetic microRNAs stimulate simultaneous, efficient, and localized regulation of multiple targets in diverse species
- PMID: 16603651
- PMCID: PMC1456869
- DOI: 10.1105/tpc.105.040725
Endogenous and synthetic microRNAs stimulate simultaneous, efficient, and localized regulation of multiple targets in diverse species
Abstract
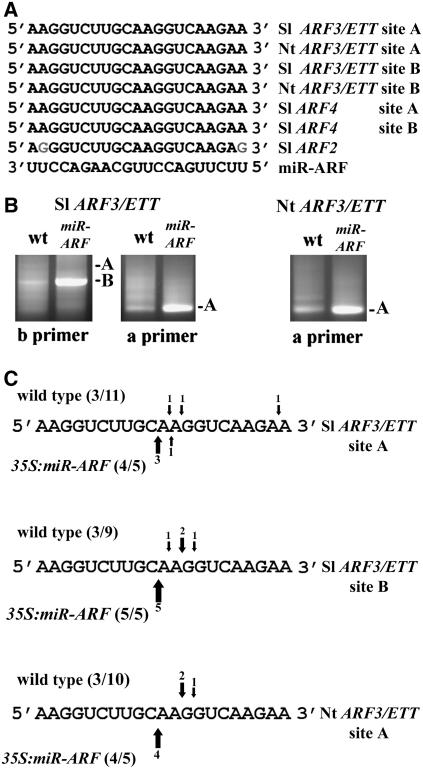
Recent studies demonstrated that pattern formation in plants involves regulation of transcription factor families by microRNAs (miRNAs). To explore the potency, autonomy, target range, and functional conservation of miRNA genes, a systematic comparison between plants ectopically expressing pre-miRNAs and plants with corresponding multiple mutant combinations of target genes was performed. We show that regulated expression of several Arabidopsis thaliana pre-miRNA genes induced a range of phenotypic alterations, the most extreme ones being a phenocopy of combined loss of their predicted target genes. This result indicates quantitative regulation by miRNA as a potential source for diversity in developmental outcomes. Remarkably, custom-made, synthetic miRNAs vectored by endogenous pre-miRNA backbones also produced phenocopies of multiple mutant combinations of genes that are not naturally regulated by miRNA. Arabidopsis-based endogenous and synthetic pre-miRNAs were also processed effectively in tomato (Solanum lycopersicum) and tobacco (Nicotiana tabacum). Synthetic miR-ARF targeting Auxin Response Factors 2, 3, and 4 induced dramatic transformations of abaxial tissues into adaxial ones in all three species, which could not cross graft joints. Likewise, organ-specific expression of miR165b that coregulates the PHABULOSA-like adaxial identity genes induced localized abaxial transformations. Thus, miRNAs provide a flexible, quantitative, and autonomous platform that can be employed for regulated expression of multiple related genes in diverse species.
Figures





References
-
- Achard, P., Herr, A., Baulcombe, D.C., and Harberd, N.P. (2004). Modulation of floral development by a gibberellin-regulated microRNA. Development 131 3357–3365. - PubMed
-
- Allen, E., Xie, Z., Gustafson, A.M., and Carrington, J.C. (2005). microRNA-directed phasing during trans-acting siRNA biogenesis in plants. Cell 121 207–221. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
- Actions
- Actions
- Actions
- Actions
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
