

Gamma-tubulin is essential for acentrosomal microtubule nucleation and coordination of late mitotic events in Arabidopsis
- PMID: 16603653
- PMCID: PMC1456865
- DOI: 10.1105/tpc.105.038364
Gamma-tubulin is essential for acentrosomal microtubule nucleation and coordination of late mitotic events in Arabidopsis
Abstract
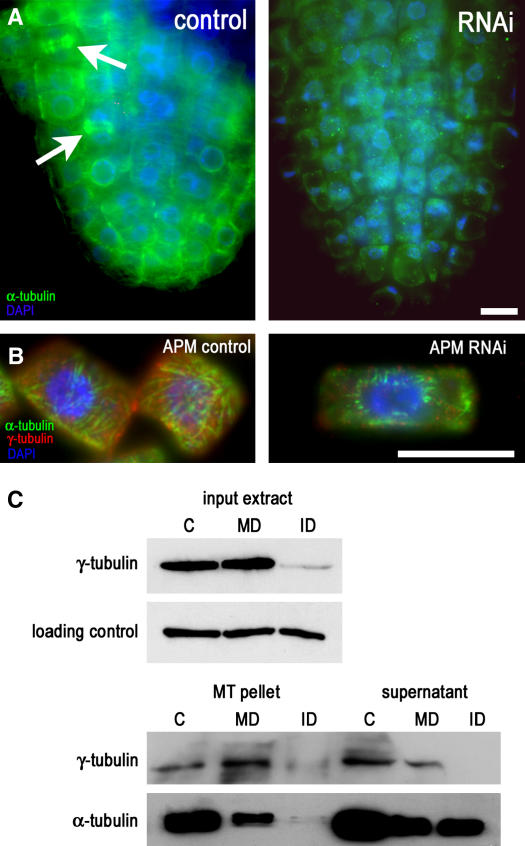
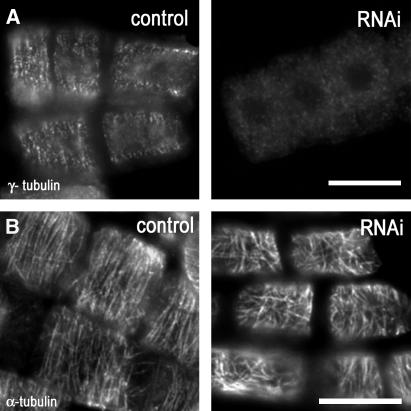
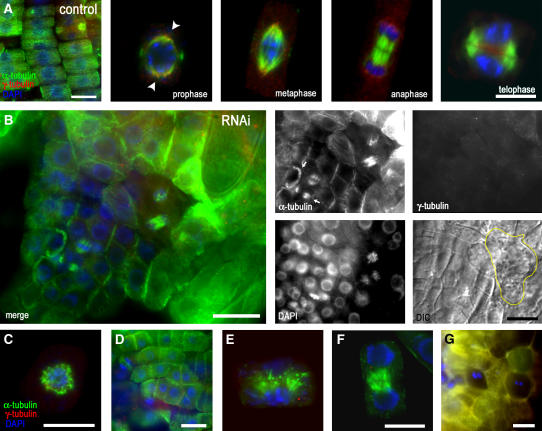
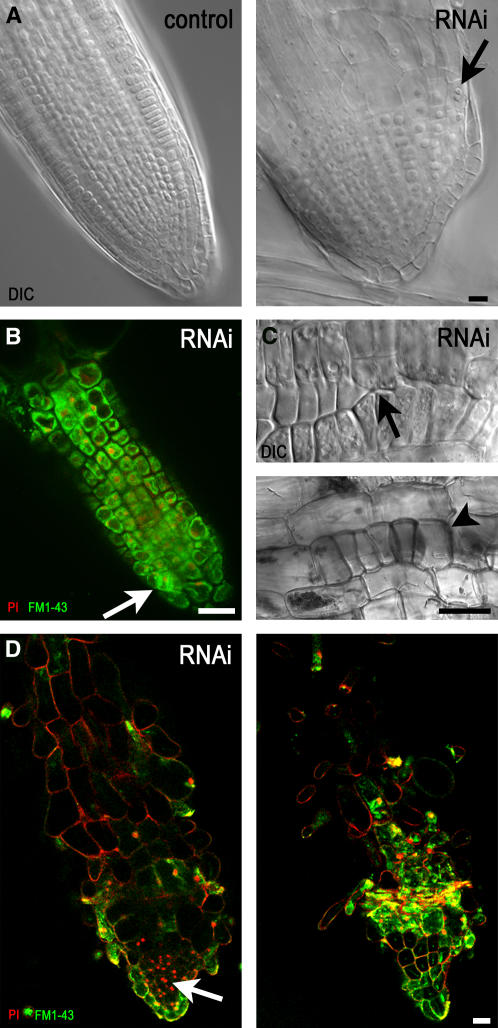
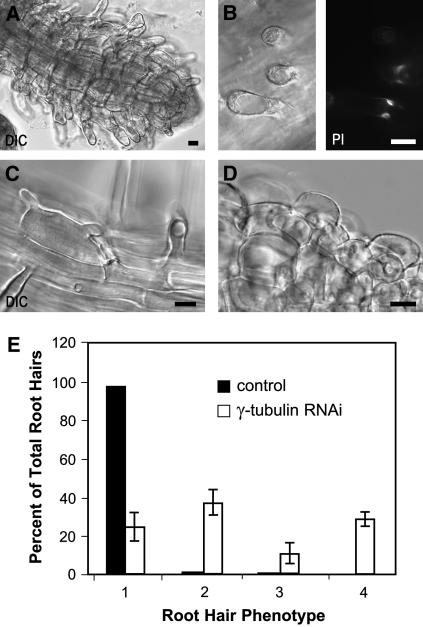
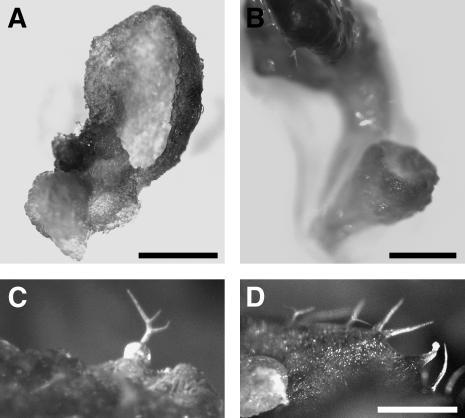
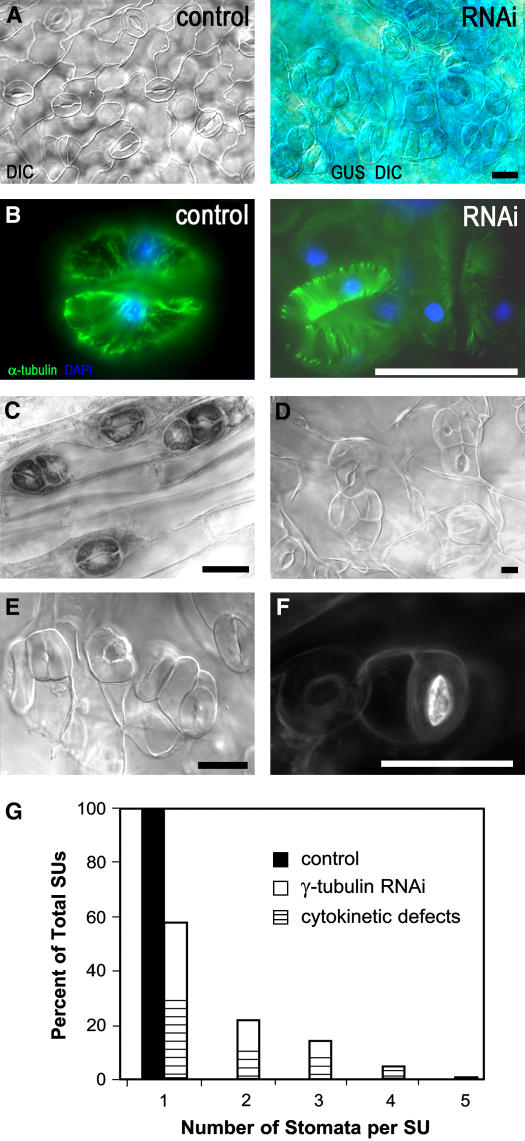
Gamma-tubulin is required for microtubule (MT) nucleation at MT organizing centers such as centrosomes or spindle pole bodies, but little is known about its noncentrosomal functions. We conditionally downregulated gamma-tubulin by inducible expression of RNA interference (RNAi) constructs in Arabidopsis thaliana. Almost complete RNAi depletion of gamma-tubulin led to the absence of MTs and was lethal at the cotyledon stage. After induction of RNAi expression, gamma-tubulin was gradually depleted from both cytoplasmic and microsomal fractions. In RNAi plants with partial loss of gamma-tubulin, MT recovery after drug-induced depolymerization was impaired. Similarly, immunodepletion of gamma-tubulin from Arabidopsis extracts severely compromised in vitro polymerization of MTs. Reduction of gamma-tubulin protein levels led to randomization and bundling of cortical MTs. This finding indicates that MT-bound gamma-tubulin is part of a cortical template guiding the microtubular network and is essential for MT nucleation. Furthermore, we found that cells with decreased levels of gamma-tubulin could progress through mitosis, but cytokinesis was strongly affected. Stepwise diminution of gamma-tubulin allowed us to reveal roles for MT nucleation in plant development, such as organization of cell files, anisotropic and polar tip growth, and stomatal patterning. Some of these functions of gamma-tubulin might be independent of MT nucleation.
Figures

References
-
- Bergmann, D.C., Lukowitz, W., and Somerville, C.R. (2004). Stomata development and pattern controlled by a MAPKK kinase. Science 304 1461–1462. - PubMed
-
- Bibikova, T.N., Blancaflor, E.B., and Gilroy, S. (1999). Microtubules regulate tip growth and orientation in root hairs of Arabidopsis thaliana. Plant J. 17 657–665. - PubMed
-
- Binarova, P., Cihalikova, J., and Dolezel, J. (1993). Localization of MPM-2 recognized phosphoproteins and tubulin during cell cycle progression in synchronized Vicia faba root meristem cells. Cell Biol. Int. 17 847–856. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
