

Microstimulation of macaque area LIP affects decision-making in a motion discrimination task
- PMID: 16604069
- PMCID: PMC2770004
- DOI: 10.1038/nn1683
Microstimulation of macaque area LIP affects decision-making in a motion discrimination task
Abstract
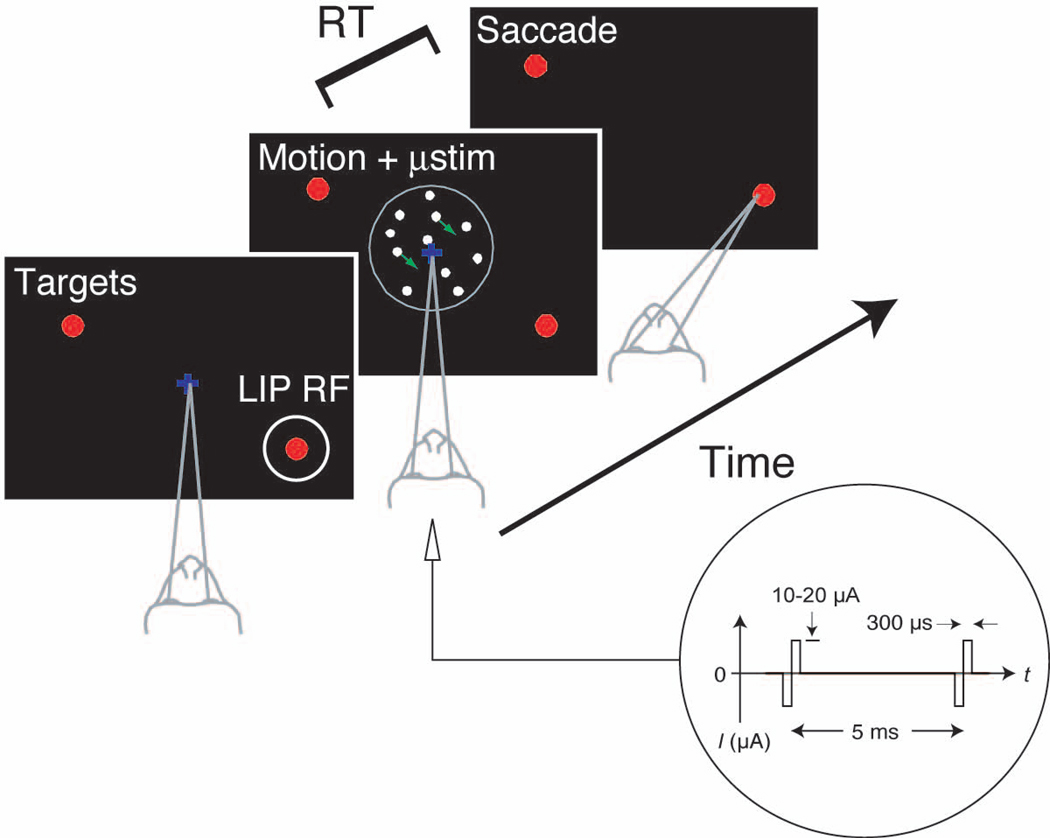
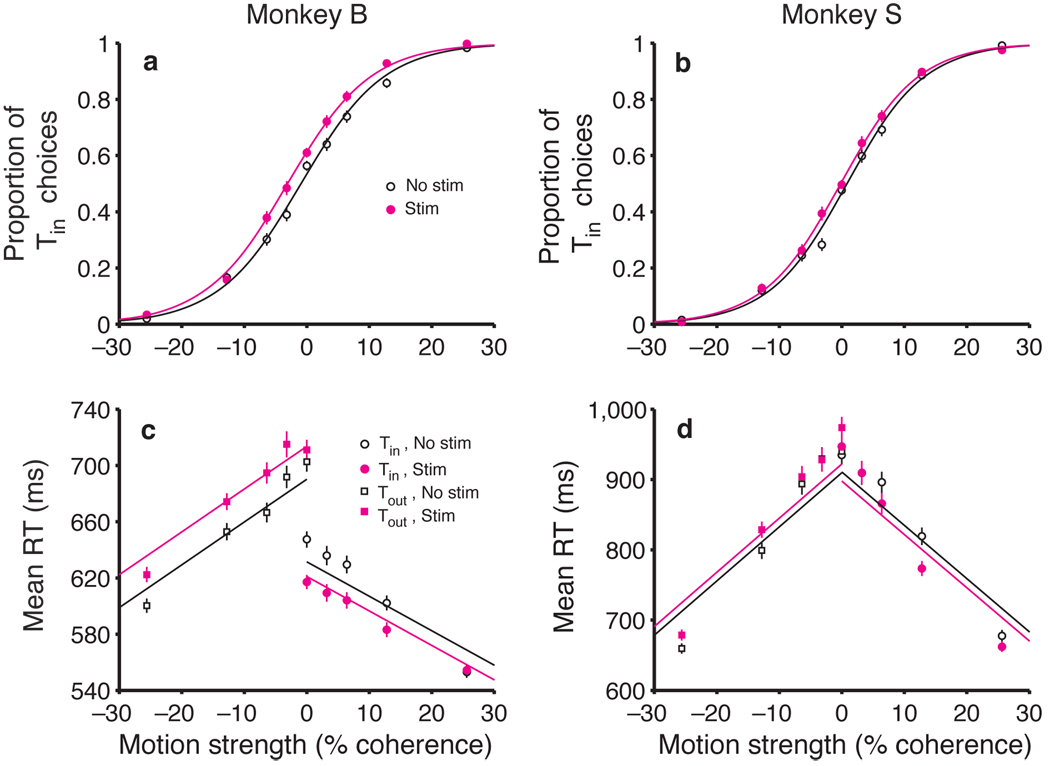
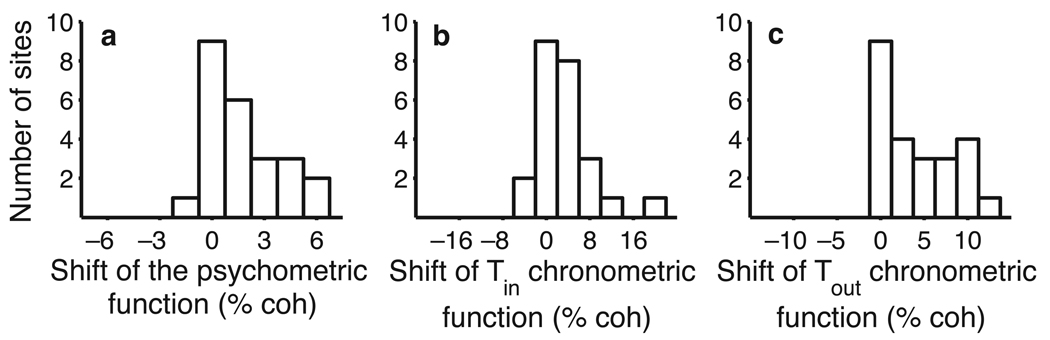
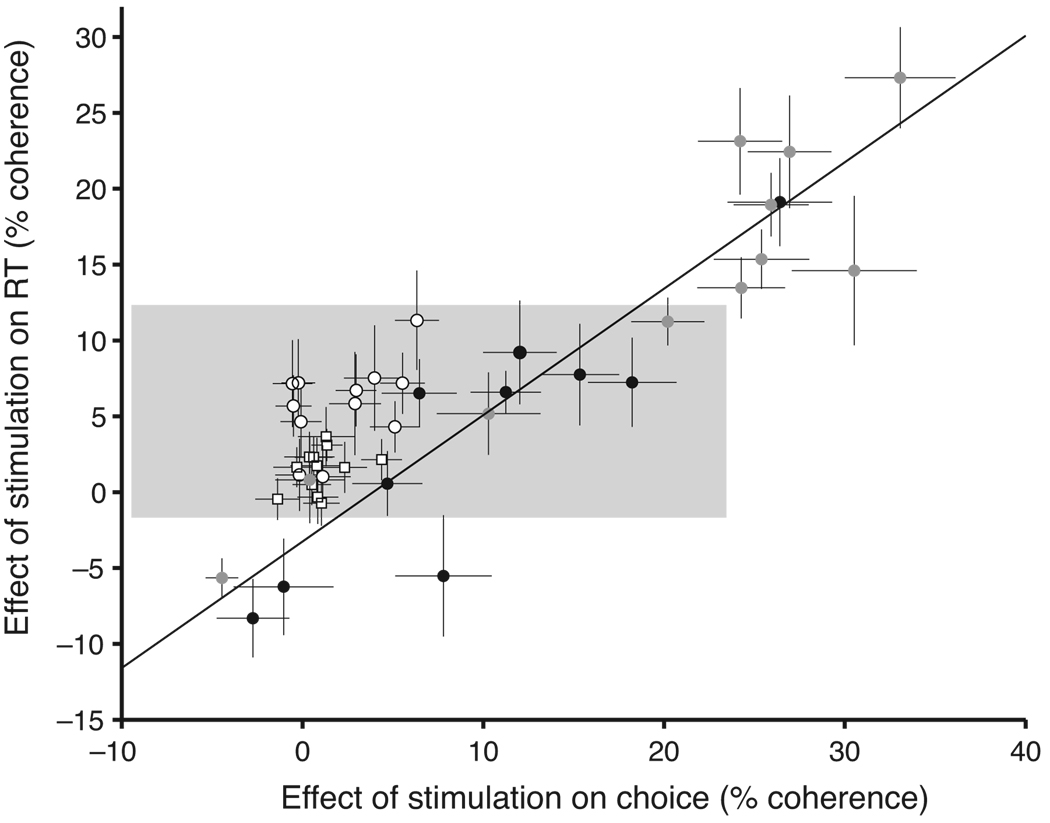
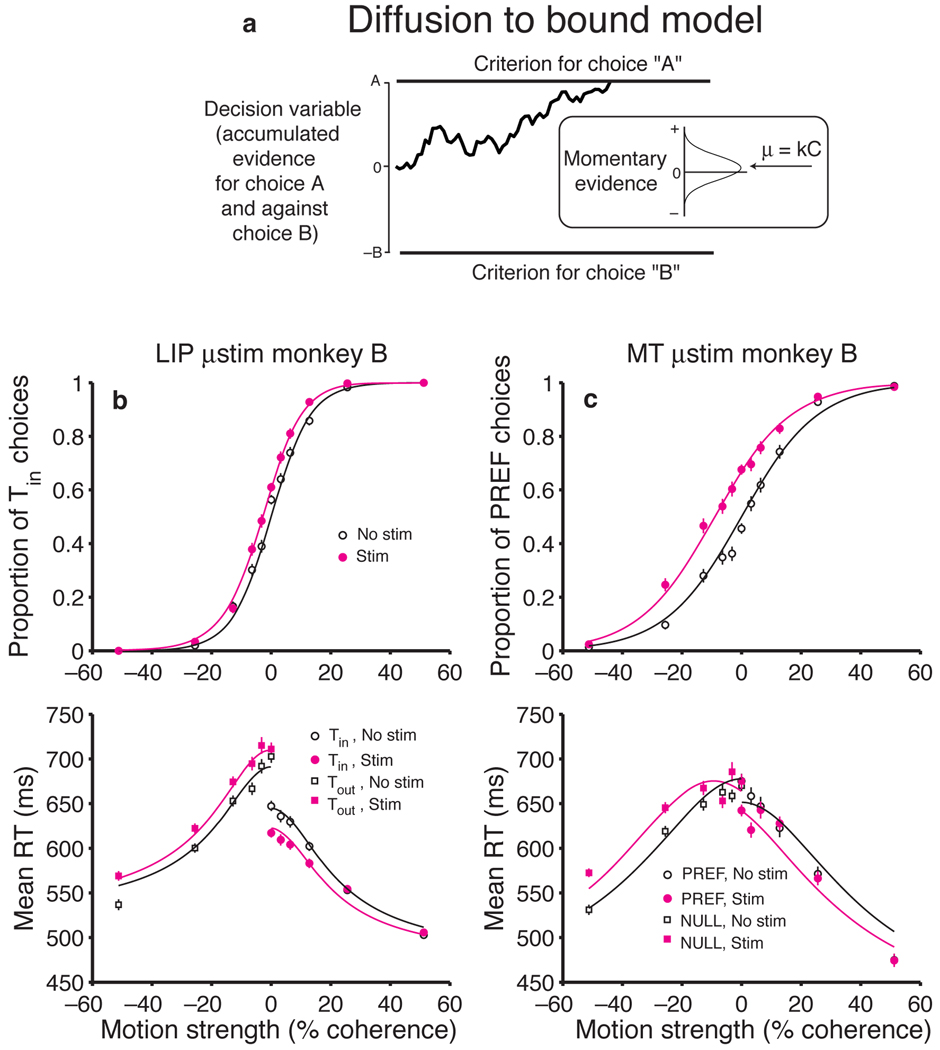
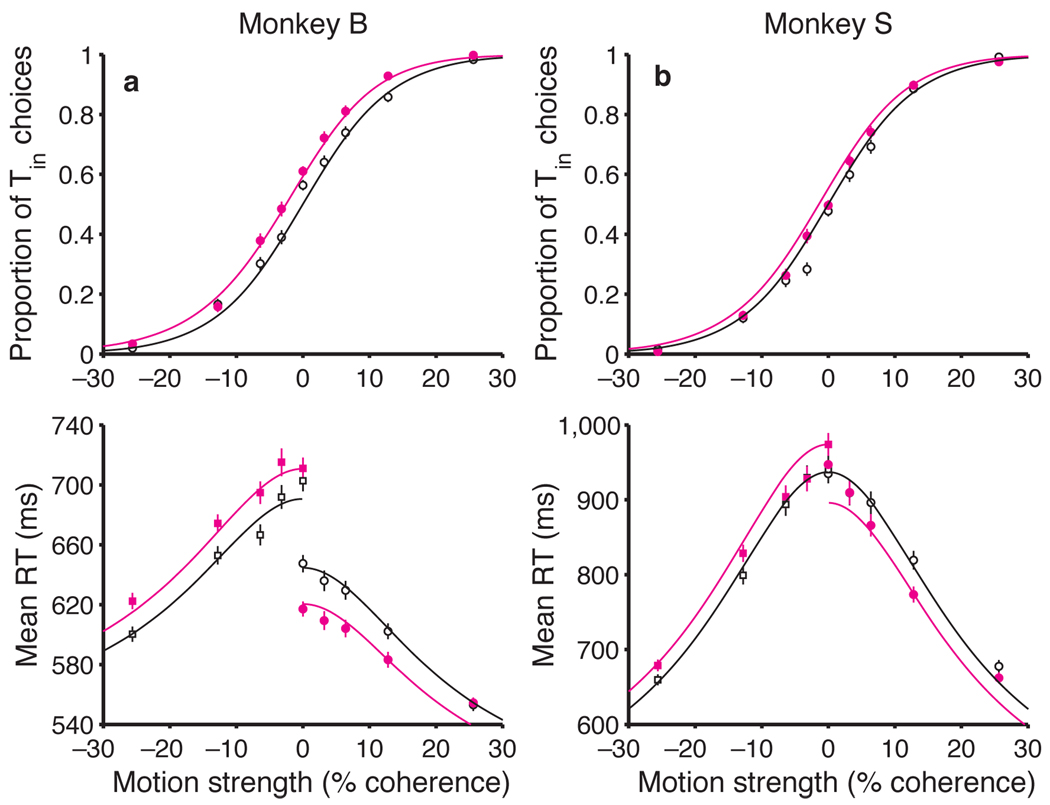
A central goal of cognitive neuroscience is to elucidate the neural mechanisms underlying decision-making. Recent physiological studies suggest that neurons in association areas may be involved in this process. To test this, we measured the effects of electrical microstimulation in the lateral intraparietal area (LIP) while monkeys performed a reaction-time motion discrimination task with a saccadic response. In each experiment, we identified a cluster of LIP cells with overlapping response fields (RFs) and sustained activity during memory-guided saccades. Microstimulation of this cluster caused an increase in the proportion of choices toward the RF of the stimulated neurons. Choices toward the stimulated RF were faster with microstimulation, while choices in the opposite direction were slower. Microstimulation never directly evoked saccades, nor did it change reaction times in a simple saccade task. These results demonstrate that the discharge of LIP neurons is causally related to decision formation in the discrimination task.
Figures
References
-
- de Lafuente V, Romo R. Neuronal correlates of subjective sensory experience. Nat Neurosci. 2005;8:1698–1703. - PubMed
-
- Schall J, Thompson K. Neural selection and control of visually guided eye movements. Annu Rev Neurosci. 1999;22:241–259. - PubMed
-
- Glimcher PW. The neurobiology of visual-saccadic decision making. Annu Rev Neurosci. 2003;26:133–179. - PubMed
-
- Romo R, Salinas E. Flutter discrimination: neural codes, perception, memory and decision making. Nat Rev Neurosci. 2003;4:203–218. - PubMed
-
- Pasternak T, Merigan WH. Motion perception following lesions of the superior temporal sulcus in the monkey. Cereb Cortex. 1994;4:247–259. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
