

Dynamic reprogramming of DNA methylation at an epigenetically sensitive allele in mice
- PMID: 16604157
- PMCID: PMC1428789
- DOI: 10.1371/journal.pgen.0020049
Dynamic reprogramming of DNA methylation at an epigenetically sensitive allele in mice
Abstract
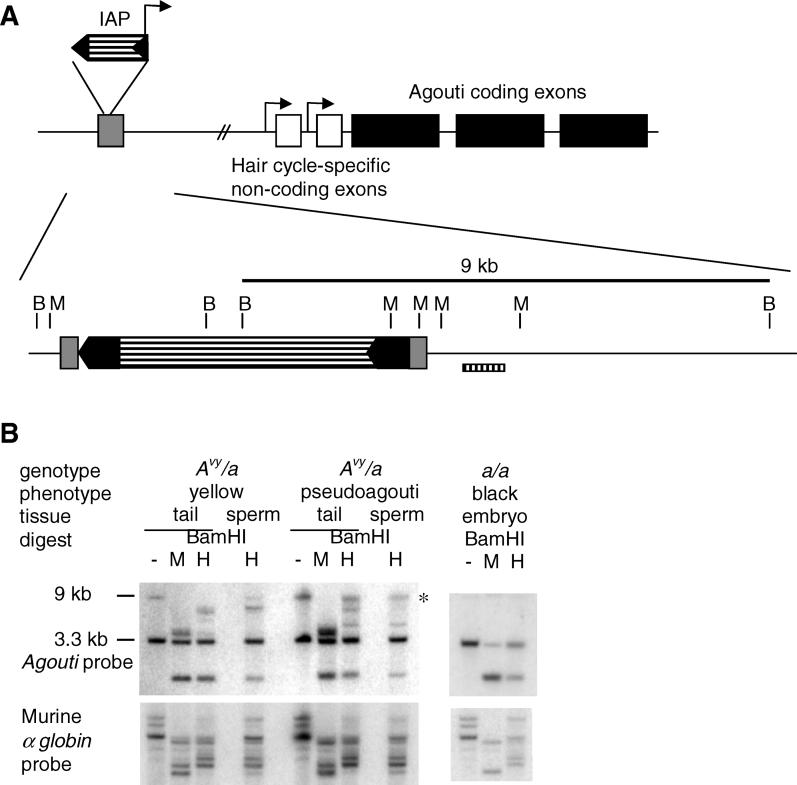
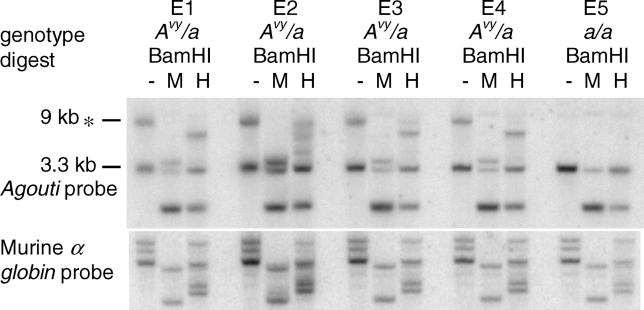
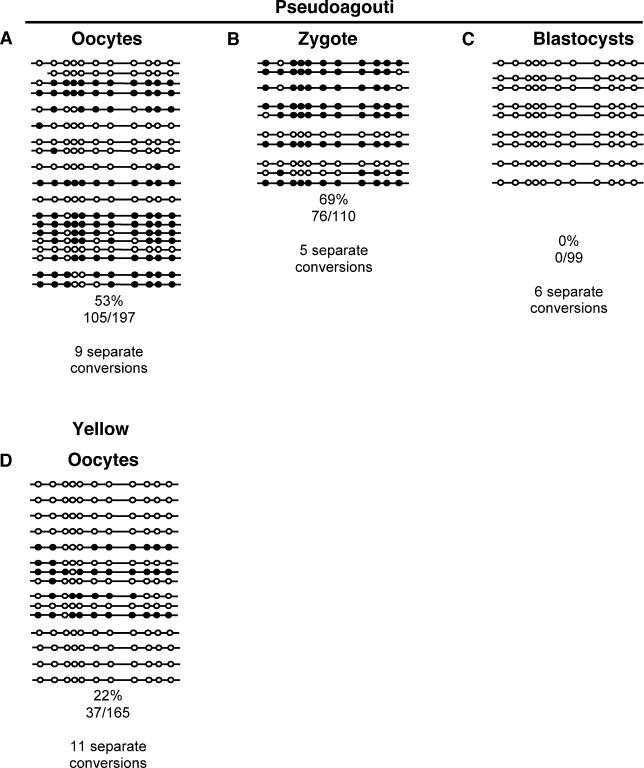
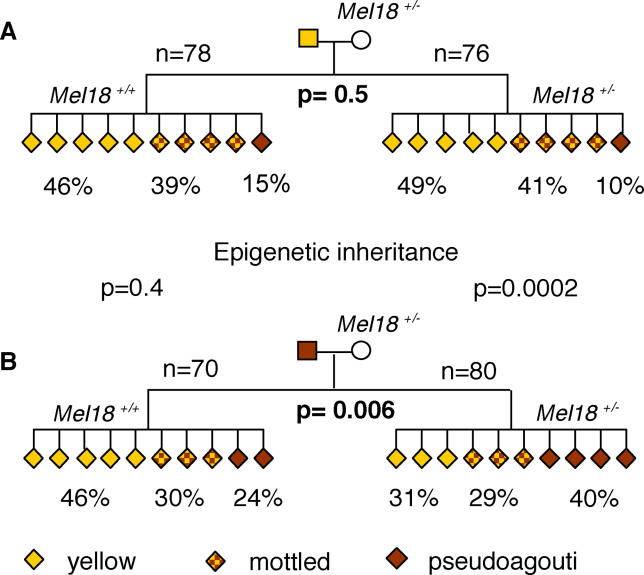
There is increasing evidence in both plants and animals that epigenetic marks are not always cleared between generations. Incomplete erasure at genes associated with a measurable phenotype results in unusual patterns of inheritance from one generation to the next, termed transgenerational epigenetic inheritance. The Agouti viable yellow (A(vy)) allele is the best-studied example of this phenomenon in mice. The A(vy) allele is the result of a retrotransposon insertion upstream of the Agouti gene. Expression at this locus is controlled by the long terminal repeat (LTR) of the retrotransposon, and expression results in a yellow coat and correlates with hypomethylation of the LTR. Isogenic mice display variable expressivity, resulting in mice with a range of coat colours, from yellow through to agouti. Agouti mice have a methylated LTR. The locus displays epigenetic inheritance following maternal but not paternal transmission; yellow mothers produce more yellow offspring than agouti mothers. We have analysed the DNA methylation in mature gametes, zygotes, and blastocysts and found that the paternally and maternally inherited alleles are treated differently. The paternally inherited allele is demethylated rapidly, and the maternal allele is demethylated more slowly, in a manner similar to that of nonimprinted single-copy genes. Interestingly, following maternal transmission of the allele, there is no DNA methylation in the blastocyst, suggesting that DNA methylation is not the inherited mark. We have independent support for this conclusion from studies that do not involve direct analysis of DNA methylation. Haplo-insufficiency for Mel18, a polycomb group protein, introduces epigenetic inheritance at a paternally derived A(vy) allele, and the pedigrees reveal that this occurs after zygotic genome activation and, therefore, despite the rapid demethylation of the locus.
Conflict of interest statement
Competing interests. The authors have declared that no competing interests exist.
Figures

References
-
- Morgan HD, Santos F, Green K, Dean W, Reik W. Epigenetic reprogramming in mammals. Hum Mol Genet 14 (Spec 1): R47–R58. 2005. - PubMed
-
- Chong S, Whitelaw E. Epigenetic germline inheritance. Curr Opin Genet Dev. 2004;14:692–696. - PubMed
-
- Morgan HD, Sutherland HG, Martin DI, Whitelaw E. Epigenetic inheritance at the agouti locus in the mouse. Nat Genet. 1999;23:314–318. - PubMed
-
- Lane N, Dean W, Erhardt S, Hajkova P, Surani A, et al. Resistance of IAPs to methylation reprogramming may provide a mechanism for epigenetic inheritance in the mouse. Genesis. 2003;35:88–93. - PubMed
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
