

Drosophila melanogaster Scramblases modulate synaptic transmission
- PMID: 16606691
- PMCID: PMC2063791
- DOI: 10.1083/jcb.200506159
Drosophila melanogaster Scramblases modulate synaptic transmission
Abstract
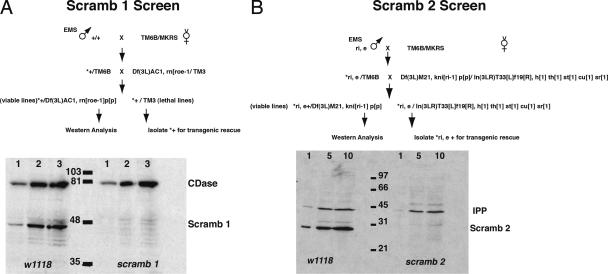
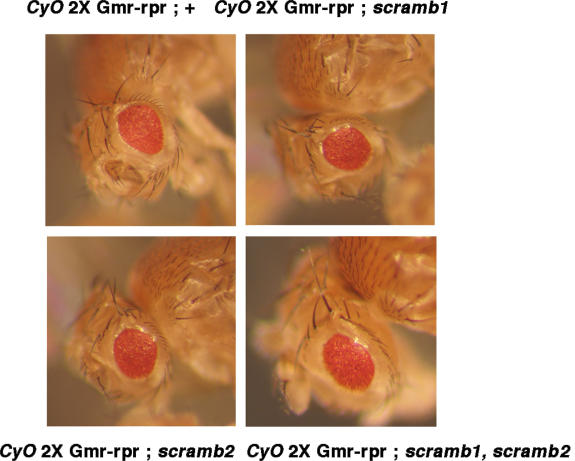
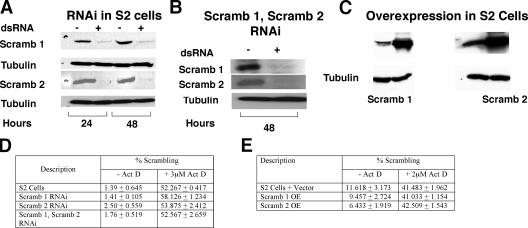
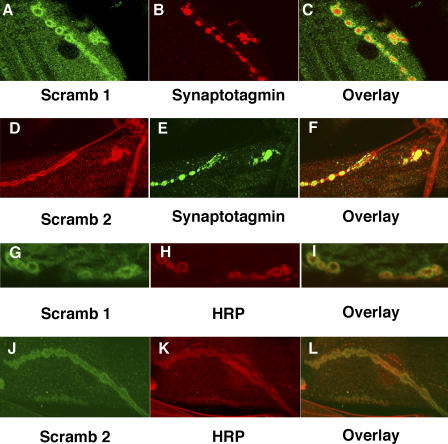
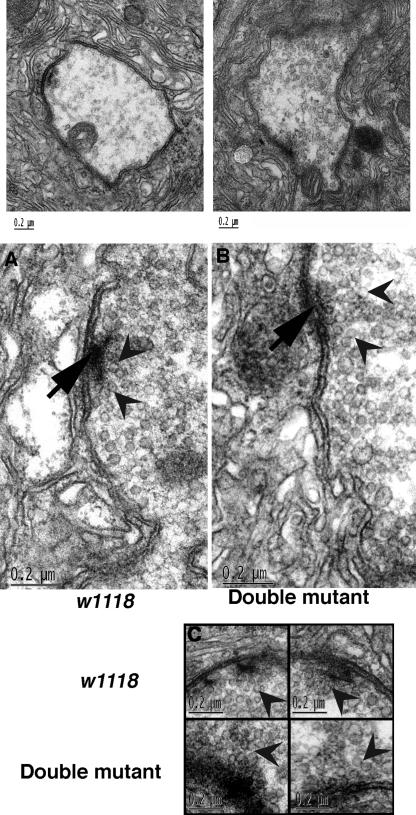
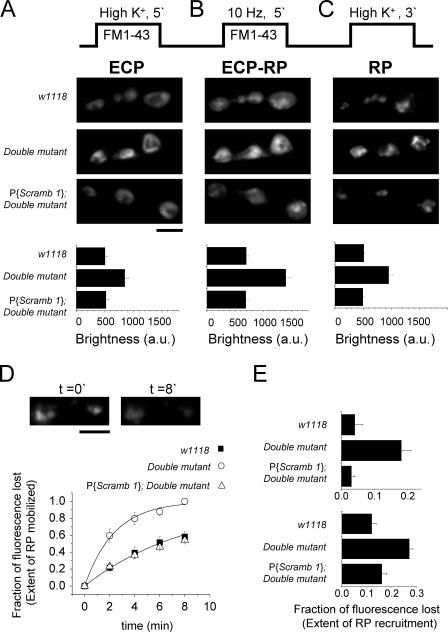
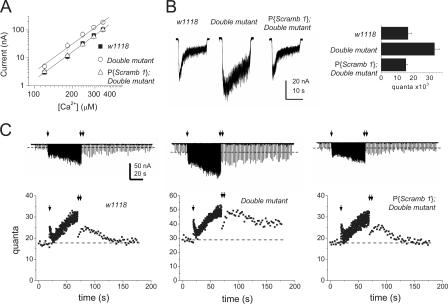
Scramblases are a family of single-pass plasma membrane proteins, identified by their purported ability to scramble phospholipids across the two layers of plasma membrane isolated from platelets and red blood cells. However, their true in vivo role has yet to be elucidated. We report the generation and isolation of null mutants of two Scramblases identified in Drosophila melanogaster. We demonstrate that flies lacking either or both of these Scramblases are not compromised in vivo in processes requiring scrambling of phospholipids. Instead, we show that D. melanogaster lacking both Scramblases have more vesicles and display enhanced recruitment from a reserve pool of vesicles and increased neurotransmitter secretion at the larval neuromuscular synapses. These defects are corrected by the introduction of a genomic copy of the Scramb 1 gene. The lack of phenotypes related to failure of scrambling and the neurophysiological analysis lead us to propose that Scramblases play a modulatory role in the process of neurotransmission.
Figures

References
-
- Acharya, J.K., P. Labarca, R. Delgado, K. Jalink, and C.S. Zuker. 1998. Synaptic defects and compensatory regulation of inositol metabolism in inositol polyphosphate 1-phosphatase mutants. Neuron. 20:1219–1229. - PubMed
-
- Acharya, U., S. Patel, E. Koundakjian, K. Nagashima, X. Han, and J.K. Acharya. 2003. Modulating sphingolipid biosynthetic pathway rescues photoreceptor degeneration. Science. 299:1740–1743. - PubMed
-
- Aho, S., and J. Uitto. 1998. Two-hybrid analysis reveals multiple direct interactions for thrombospondin 1. Matrix Biol. 17:401–412. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
