

Haplotype variation in structure and expression of a gene cluster associated with a quantitative trait locus for improved yield in rice
- PMID: 16606701
- PMCID: PMC1457051
- DOI: 10.1101/gr.4814006
Haplotype variation in structure and expression of a gene cluster associated with a quantitative trait locus for improved yield in rice
Abstract
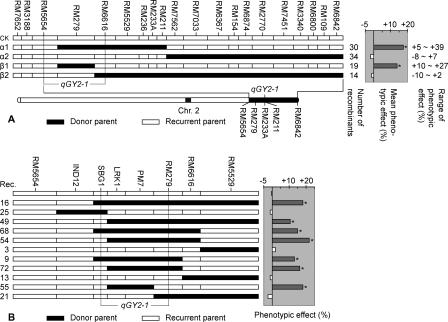
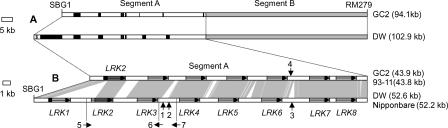
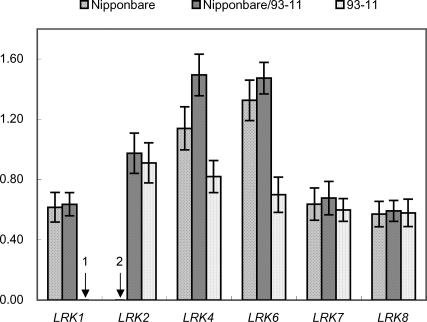
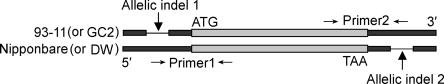
By constructing nearly isogenic lines (NILs) that differ only at a single quantitative trait locus (QTL), we fine-mapped the yield-improving QTL qGY2-1 to a 102.9-kb region on rice chromosome 2. Comparison analysis of the genomic sequences in the mapped QTL region between the donor (Dongxiang wild rice, Oryza rufipogon Griff.) and recurrent (Guichao2, Oryza sativa ssp. indica) parents used for the development of NILs identified the haplotypes of a leucine-rich repeat receptor kinase gene cluster, which showed extensive allelic variation. The sequences between genes in the cluster had a very high rate of divergence. More importantly, the genes themselves also differed between two haplotypes: Only 92% identity was observed for one allele, and another allele was found to have completely lost its allelic counterpart in Guichao2. The other six shared genes all showed >98% identity, and four of these exhibited obvious regulatory variation. The same haplotype segments also differed in length (43.9-kb in Guichao2 vs. 52.6-kb in Dongxiang wild rice). Such extensive sequence variation was also observed between orthologous regions of indica (cv. 93-11) and japonica (cv. Nipponbare) subspecies of Oryza sativa. Different rates of sequence divergence within the cluster have resulted in haplotype variability in 13 rice accessions. We also detected allelic expression variation in this gene cluster, in which some genes gave unequal expression of alleles in hybrids. These allelic variations in structure and expression suggest that the leucine-rich repeat receptor kinase gene cluster identified in our study should be a particularly good candidate for the source of the yield QTL.
Figures

References
-
- Comeron J.M. K-Estimator: Calculation of the number of nucleotide substitutions per site and the confidence intervals. Bioinformatics. 1999;15:763–764. - PubMed
-
- Cowles C.R., Hirschhorn J.N., Altshuler D., Lander E.S., Hirschhorn J.N., Altshuler D., Lander E.S., Altshuler D., Lander E.S., Lander E.S. Detection of regulatory variation in mouse genes. Nat. Genet. 2002;32:432–437. - PubMed
-
- Doebley J., Stec A., Hubbard L., Stec A., Hubbard L., Hubbard L. The evolution of apical dominance in maize. Nature. 1997;386:485–488. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources