

Evolutionary expansion and divergence in the ZNF91 subfamily of primate-specific zinc finger genes
- PMID: 16606703
- PMCID: PMC1457049
- DOI: 10.1101/gr.4843906
Evolutionary expansion and divergence in the ZNF91 subfamily of primate-specific zinc finger genes
Abstract
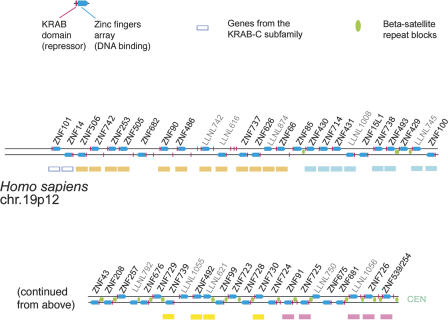
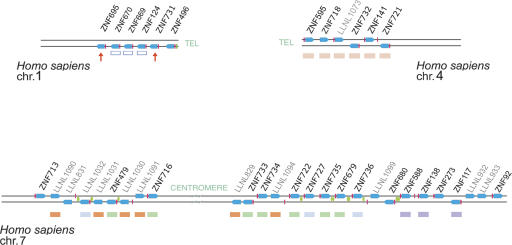
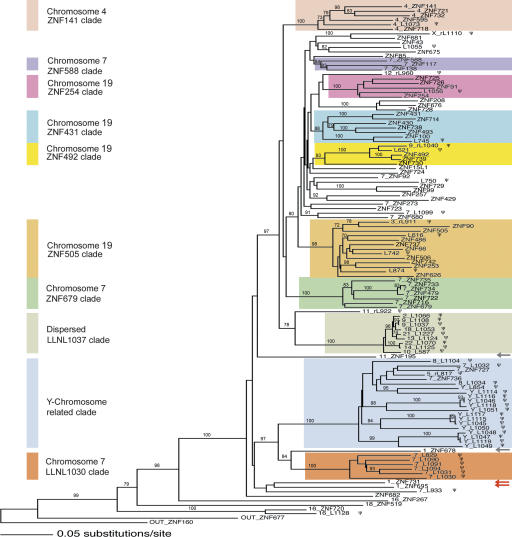
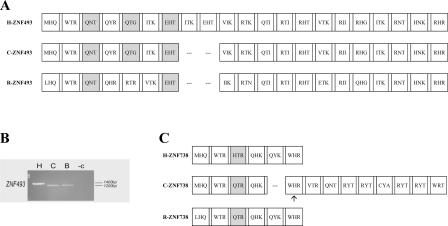
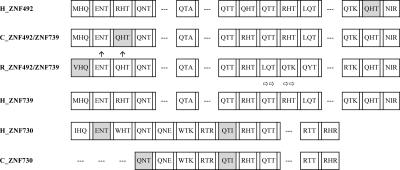
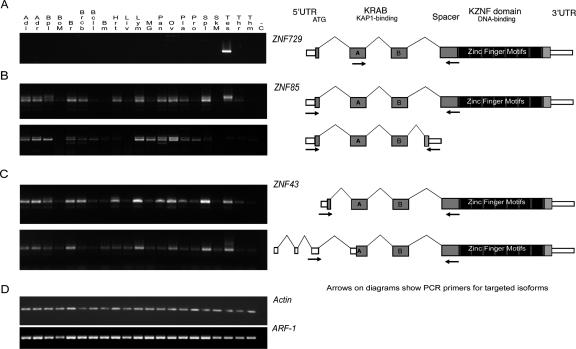
Most genes are conserved in mammals, but certain gene families have acquired large numbers of lineage-specific loci through repeated rounds of gene duplication, divergence, and loss that have continued in each mammalian group. One such family encodes KRAB-zinc finger (KRAB-ZNF) proteins, which function as transcriptional repressors. One particular subfamily of KRAB-ZNF genes, including ZNF91, has expanded specifically in primates to comprise more than 110 loci in the human genome. Genes of the ZNF91 subfamily reside in large gene clusters near centromeric regions of human chromosomes 19 and 7 with smaller clusters or isolated copies in other locations. Phylogenetic analysis indicates that many of these genes arose before the split between the New and Old World monkeys, but the ZNF91 subfamily has continued to expand and diversify throughout the evolution of apes and humans. Paralogous loci are distinguished by divergence within their zinc finger arrays, indicating selection for proteins with different regulatory targets. In addition, many loci produce multiple alternatively spliced transcripts encoding proteins that may serve separate and perhaps even opposing regulatory roles because of the modular motif structure of KRAB-ZNF genes. The tissue-specific expression patterns and rapid structural divergence of ZNF91 subfamily genes suggest a role in determining gene expression differences between species and the evolution of novel primate traits.
Figures

Similar articles
-
A comprehensive catalog of human KRAB-associated zinc finger genes: insights into the evolutionary history of a large family of transcriptional repressors.Genome Res. 2006 May;16(5):669-77. doi: 10.1101/gr.4842106. Epub 2006 Apr 10. Genome Res. 2006. PMID: 16606702 Free PMC article.
-
Evolution of C2H2-zinc finger genes and subfamilies in mammals: species-specific duplication and loss of clusters, genes and effector domains.BMC Evol Biol. 2008 Jun 18;8:176. doi: 10.1186/1471-2148-8-176. BMC Evol Biol. 2008. PMID: 18559114 Free PMC article.
-
Emergence of the ZNF91 Krüppel-associated box-containing zinc finger gene family in the last common ancestor of anthropoidea.Proc Natl Acad Sci U S A. 1995 Nov 7;92(23):10757-61. doi: 10.1073/pnas.92.23.10757. Proc Natl Acad Sci U S A. 1995. PMID: 7479878 Free PMC article.
-
Evolution of KRAB-containing zinc finger proteins and their roles in species evolution.Yi Chuan. 2016 Nov 20;38(11):971-978. doi: 10.16288/j.yczz.16-056. Yi Chuan. 2016. PMID: 27867147 Review.
-
Lineage-specific expansion of KRAB zinc-finger transcription factor genes: implications for the evolution of vertebrate regulatory networks.Cold Spring Harb Symp Quant Biol. 2003;68:131-40. doi: 10.1101/sqb.2003.68.131. Cold Spring Harb Symp Quant Biol. 2003. PMID: 15338611 Review. No abstract available.
Cited by
-
Epigenetic regulation of puberty via Zinc finger protein-mediated transcriptional repression.Nat Commun. 2015 Dec 16;6:10195. doi: 10.1038/ncomms10195. Nat Commun. 2015. PMID: 26671628 Free PMC article.
-
Zinc-finger domains in metazoans: evolution gone wild.Genome Biol. 2017 Sep 6;18(1):168. doi: 10.1186/s13059-017-1307-y. Genome Biol. 2017. PMID: 28877710 Free PMC article. No abstract available.
-
ZNF-mediated resistance to imatinib mesylate in gastrointestinal stromal tumor.PLoS One. 2013;8(1):e54477. doi: 10.1371/journal.pone.0054477. Epub 2013 Jan 25. PLoS One. 2013. PMID: 23372733 Free PMC article.
-
Accumulation of interspersed and sex-specific repeats in the non-recombining region of papaya sex chromosomes.BMC Genomics. 2014 May 4;15(1):335. doi: 10.1186/1471-2164-15-335. BMC Genomics. 2014. PMID: 24885930 Free PMC article.
-
Hominoid SVA-lncRNA AK057321 targets human-specific SVA retrotransposons in SCN8A and CDK5RAP2 to initiate neuronal maturation.Commun Biol. 2023 Mar 30;6(1):347. doi: 10.1038/s42003-023-04683-8. Commun Biol. 2023. PMID: 36997626 Free PMC article.
References
-
- Abrink M., Larsson E., Hellman L., Larsson E., Hellman L., Hellman L. Demethylation of ERV3, an endogenous retrovirus regulating the Kruppel-related zinc finger gene H-plk, in several human cell lines arrested during early monocyte development. DNA Cell Biol. 1998;17:27–37. - PubMed
-
- Ayyanathan K., Lechner M.S., Bell P., Maul G.G., Schultz D.C., Yamada Y., Tanaka K., Torigoe K., Rauscher 3rd F.J., Lechner M.S., Bell P., Maul G.G., Schultz D.C., Yamada Y., Tanaka K., Torigoe K., Rauscher 3rd F.J., Bell P., Maul G.G., Schultz D.C., Yamada Y., Tanaka K., Torigoe K., Rauscher 3rd F.J., Maul G.G., Schultz D.C., Yamada Y., Tanaka K., Torigoe K., Rauscher 3rd F.J., Schultz D.C., Yamada Y., Tanaka K., Torigoe K., Rauscher 3rd F.J., Yamada Y., Tanaka K., Torigoe K., Rauscher 3rd F.J., Tanaka K., Torigoe K., Rauscher 3rd F.J., Torigoe K., Rauscher 3rd F.J., Rauscher 3rd F.J. Regulated recruitment of HP1 to a euchromatic gene induces mitotically heritable, epigenetic gene silencing: A mammalian cell culture model of gene variegation. Genes & Dev. 2003;17:1855–1869. - PMC - PubMed
-
- Bailey J.A., Yavor A.M., Massa H.F., Trask B.J., Eichler E.E., Yavor A.M., Massa H.F., Trask B.J., Eichler E.E., Massa H.F., Trask B.J., Eichler E.E., Trask B.J., Eichler E.E., Eichler E.E. Segmental duplications: Organization and impact within the current human genome project assembly. Genome Res. 2001;11:1005–1017. - PMC - PubMed
-
- Bailey J.A., Gu Z., Clark R.A., Reinert K., Samonte R.V., Schwartz S., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Gu Z., Clark R.A., Reinert K., Samonte R.V., Schwartz S., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Clark R.A., Reinert K., Samonte R.V., Schwartz S., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Reinert K., Samonte R.V., Schwartz S., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Samonte R.V., Schwartz S., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Schwartz S., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Adams M.D., Myers E.W., Li P.W., Eichler E.E., Myers E.W., Li P.W., Eichler E.E., Li P.W., Eichler E.E., Eichler E.E. Recent segmental duplications in the human genome. Science. 2002;297:1003–1007. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials