

Hsp70 chaperones accelerate protein translocation and the unfolding of stable protein aggregates by entropic pulling
- PMID: 16606842
- PMCID: PMC1458849
- DOI: 10.1073/pnas.0510496103
Hsp70 chaperones accelerate protein translocation and the unfolding of stable protein aggregates by entropic pulling
Abstract
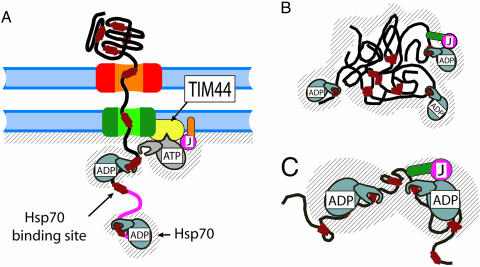
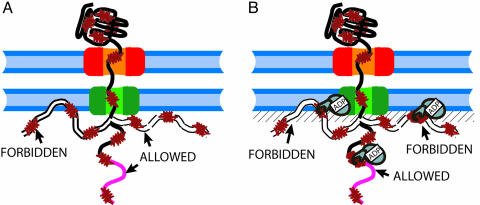
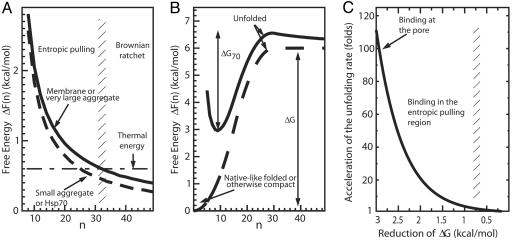
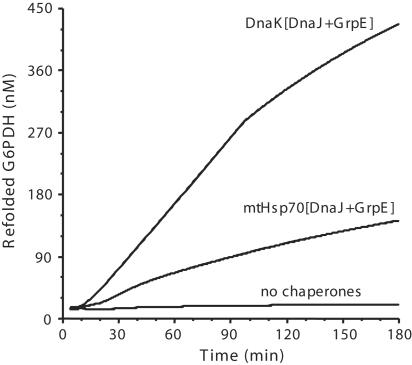
Hsp70s are highly conserved ATPase molecular chaperones mediating the correct folding of de novo synthesized proteins, the translocation of proteins across membranes, the disassembly of some native protein oligomers, and the active unfolding and disassembly of stress-induced protein aggregates. Here, we bring thermodynamic arguments and biochemical evidences for a unifying mechanism named entropic pulling, based on entropy loss due to excluded-volume effects, by which Hsp70 molecules can convert the energy of ATP hydrolysis into a force capable of accelerating the local unfolding of various protein substrates and, thus, perform disparate cellular functions. By means of entropic pulling, individual Hsp70 molecules can accelerate unfolding and pulling of translocating polypeptides into mitochondria in the absence of a molecular fulcrum, thus settling former contradictions between the power-stroke and the Brownian ratchet models for Hsp70-mediated protein translocation across membranes. Moreover, in a very different context devoid of membrane and components of the import pore, the same physical principles apply to the forceful unfolding, solubilization, and assisted native refolding of stable protein aggregates by individual Hsp70 molecules, thus providing a mechanism for Hsp70-mediated protein disaggregation.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
