

Separation of mu-opioid receptor desensitization and internalization: endogenous receptors in primary neuronal cultures
- PMID: 16611829
- PMCID: PMC6673897
- DOI: 10.1523/JNEUROSCI.0303-06.2006
Separation of mu-opioid receptor desensitization and internalization: endogenous receptors in primary neuronal cultures
Abstract
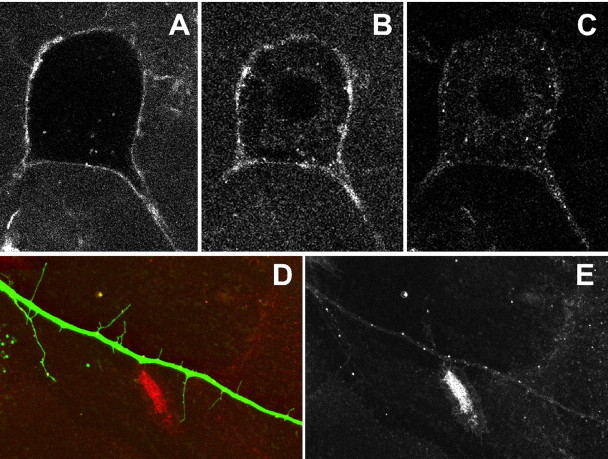
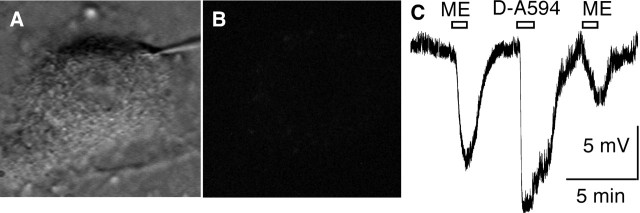
A close relationship between desensitization and internalization of mu-opioid receptors (MORs) has been proposed based on differential actions of series of agonists. The role that these two processes have in the development of tolerance and dependence to opioids has been a controversial subject that has been studied in a variety of model systems. Here, we examine desensitization and internalization of endogenous MORs simultaneously in primary cultures of locus ceruleus neurons using fluorescently tagged peptide agonists. With the use of two fluorescent opioid peptides, dermorphin-Bodipy Texas Red and dermorphin-Alexa594 (Derm-A594), desensitization was measured electrophysiologically and trafficking was followed by the accumulation of intracellular fluorescent puncta. Blocking endocytosis with concanavalin A eliminated the accumulation of fluorescent puncta but desensitization induced by Derm-A594 was unaffected. Likewise, after treatment with concanavalin A, there was no change in either desensitization or recovery from desensitization induced by [Met]5enkephalin. The results demonstrate that desensitization and the recovery from desensitization are not dependent on receptor internalization and suggest that the activity of endogenous MORs in primary neurons can be modulated at the level of the plasma membrane.
Figures






References
-
- Arttamangkul S, Alvarez-Maubecin V, Thomas G, Williams JT, Grandy DK (2000). Binding and internalization of fluorescent opioid peptide conjugates in living cells. Mol Pharmacol 58:1570–1580. - PubMed
-
- Bailey CP, Connor M (2005). Opioids: cellular mechanisms of tolerance and physical dependence. Curr Opin Pharmacol 5:60–68. - PubMed
-
- Bailey CP, Kelly E, Henderson G (2004). Protein kinase C activation enhances morphine-induced rapid desensitization of mu-opioid receptors in mature rat locus ceruleus neurons. Mol Pharmacol 66:1592–1598. - PubMed
-
- Beaumont V, Hepworth MB, Luty JS, Kelly E, Henderson G (1998). Somatostatin receptor desensitization in NG108–15 cells. A consequence of receptor sequestration. J Biol Chem 273:33174–33183. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous