

Protein kinase C delta activates topoisomerase IIalpha to induce apoptotic cell death in response to DNA damage
- PMID: 16611985
- PMCID: PMC1447416
- DOI: 10.1128/MCB.26.9.3414-3431.2006
Protein kinase C delta activates topoisomerase IIalpha to induce apoptotic cell death in response to DNA damage
Abstract
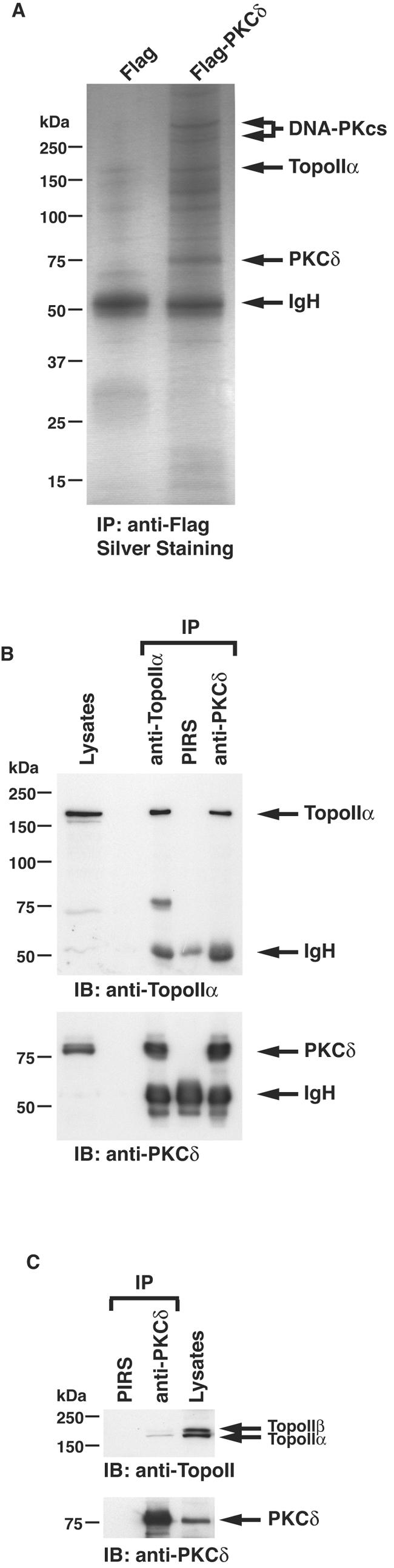
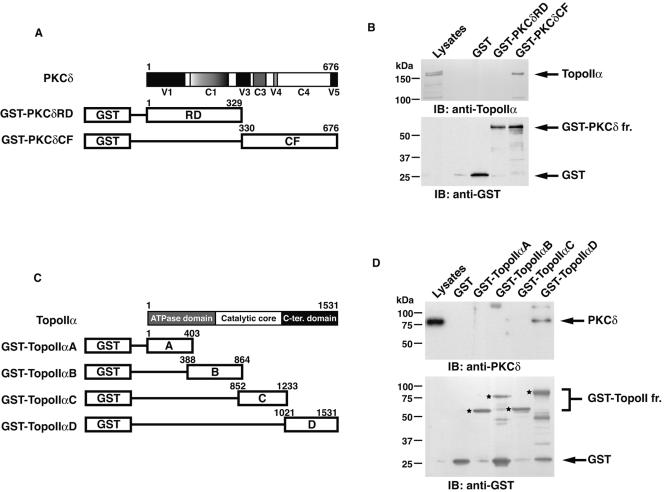
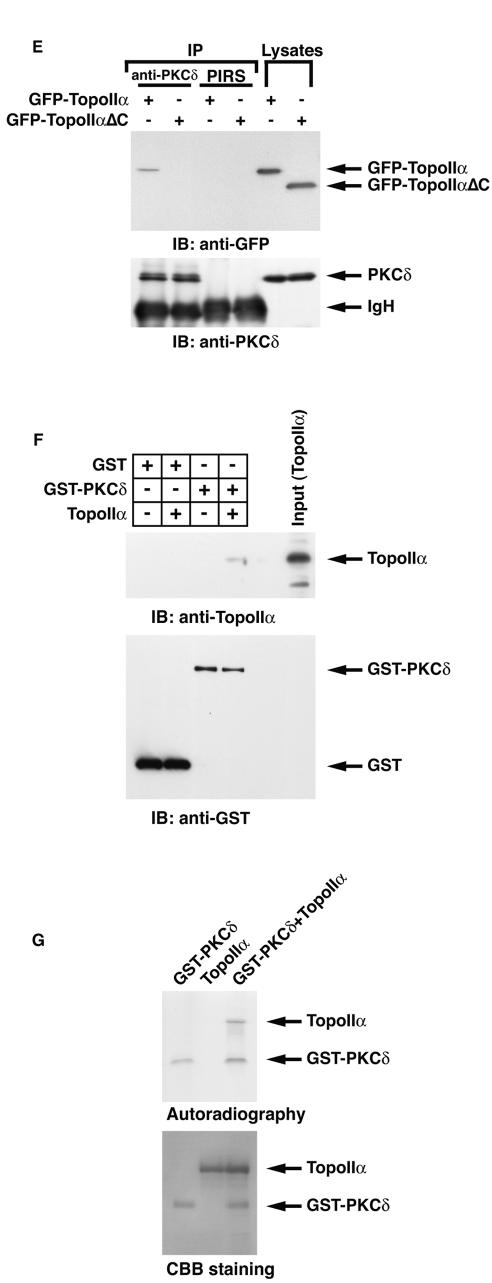
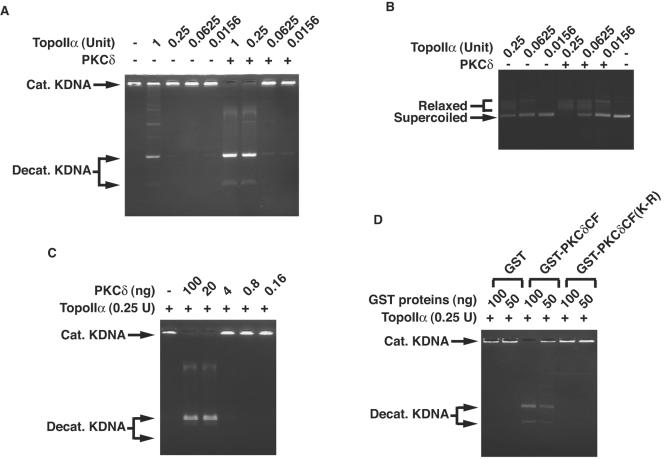
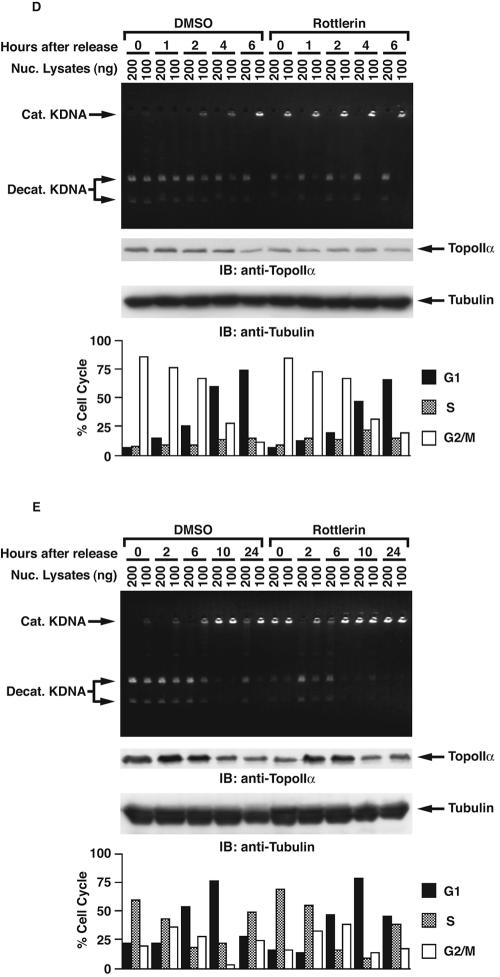
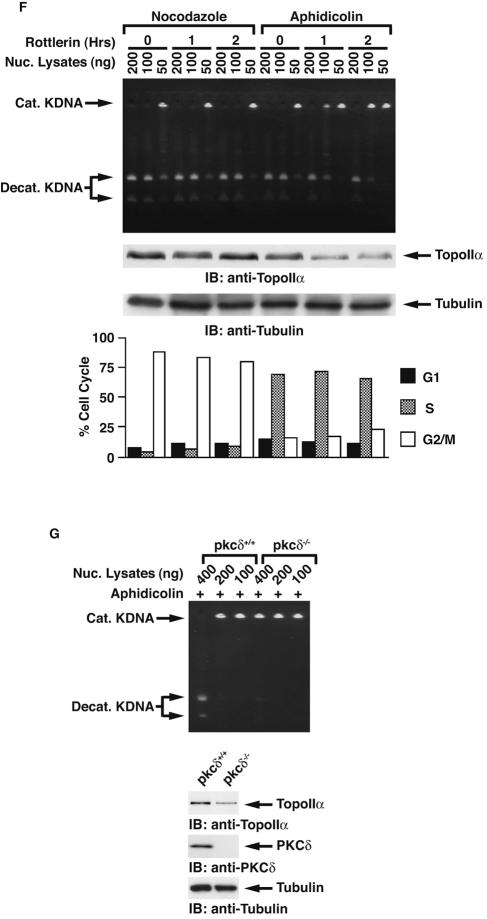
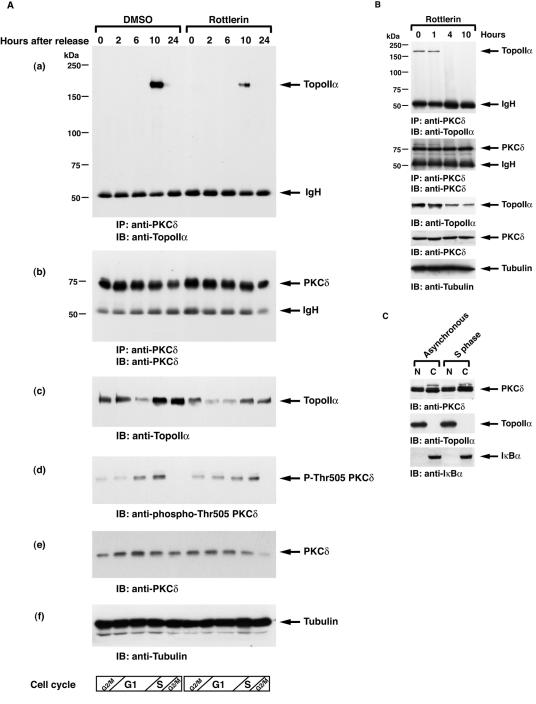
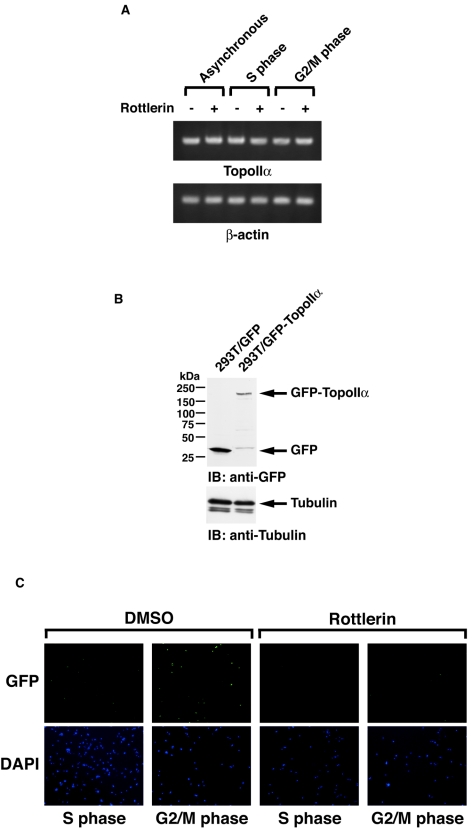
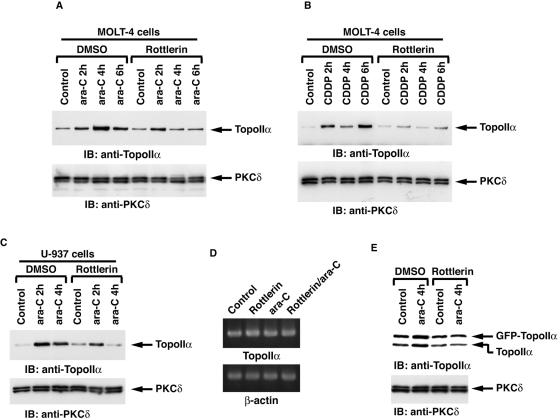
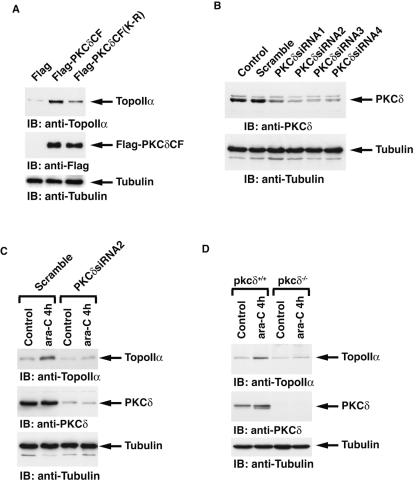
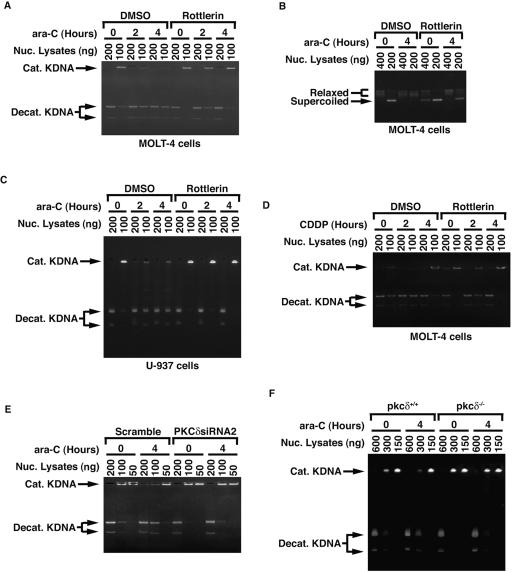
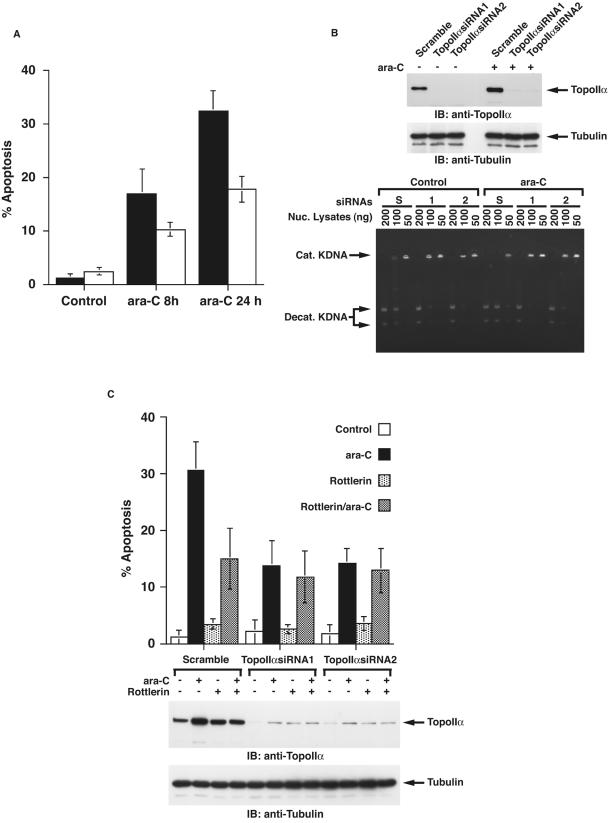
DNA topoisomerase II is an essential nuclear enzyme that modulates DNA processes by altering the topological state of double-stranded DNA. This enzyme is required for chromosome condensation and segregation; however, the regulatory mechanism of its activation is largely unknown. Here we demonstrate that topoisomerase IIalpha is activated in response to genotoxic stress. Concomitant with the activation, the expression of topoisomerase IIalpha is increased following DNA damage. The results also demonstrate that the proapoptotic kinase protein kinase C delta (PKCdelta) interacts with topoisomerase IIalpha. This association is in an S-phase-specific manner and is required for stabilization and catalytic activation of topoisomerase IIalpha in response to DNA damage. Conversely, inhibition of PKCdelta activity attenuates DNA damage-induced activation of topoisomerase IIalpha. Finally, aberrant activation of topoisomerase IIalpha by PKCdelta is associated with induction of apoptosis upon exposure to genotoxic agents. These findings indicate that PKCdelta regulates topoisomerase IIalpha and thereby cell fate in the genotoxic stress response.
Figures


Similar articles
-
The p53 tumor suppressor stimulates the catalytic activity of human topoisomerase IIalpha by enhancing the rate of ATP hydrolysis.J Biol Chem. 2000 Jun 16;275(24):18503-10. doi: 10.1074/jbc.M002081200. J Biol Chem. 2000. PMID: 10764786
-
HU-331 is a catalytic inhibitor of topoisomerase IIα.Chem Res Toxicol. 2014 Dec 15;27(12):2044-51. doi: 10.1021/tx500245m. Epub 2014 Dec 3. Chem Res Toxicol. 2014. PMID: 25409338
-
Fbxo28 promotes mitotic progression and regulates topoisomerase IIα-dependent DNA decatenation.Cell Cycle. 2016 Dec 16;15(24):3419-3431. doi: 10.1080/15384101.2016.1246093. Epub 2016 Oct 18. Cell Cycle. 2016. PMID: 27754753 Free PMC article.
-
PKCdelta signaling: mechanisms of DNA damage response and apoptosis.Cell Signal. 2007 May;19(5):892-901. doi: 10.1016/j.cellsig.2007.01.027. Epub 2007 Feb 7. Cell Signal. 2007. PMID: 17336499 Review.
-
Recent advances in the development of catalytic inhibitors of human DNA topoisomerase IIα as novel anticancer agents.Curr Med Chem. 2013;20(5):694-709. doi: 10.2174/092986713804999402. Curr Med Chem. 2013. PMID: 23210851 Review.
Cited by
-
Molecular markers in oral lichen planus: A systematic review.J Oral Maxillofac Pathol. 2016 Jan-Apr;20(1):115-21. doi: 10.4103/0973-029X.180964. J Oral Maxillofac Pathol. 2016. PMID: 27194873 Free PMC article. Review.
-
ATM augments nuclear stabilization of DYRK2 by inhibiting MDM2 in the apoptotic response to DNA damage.J Biol Chem. 2010 Feb 12;285(7):4909-19. doi: 10.1074/jbc.M109.042341. Epub 2009 Dec 4. J Biol Chem. 2010. PMID: 19965871 Free PMC article.
-
Inhibiting tyrosine phosphorylation of protein kinase Cδ (PKCδ) protects the salivary gland from radiation damage.J Biol Chem. 2014 Apr 11;289(15):10900-10908. doi: 10.1074/jbc.M114.551366. Epub 2014 Feb 25. J Biol Chem. 2014. PMID: 24569990 Free PMC article.
-
Doxorubicin induced heart failure: Phenotype and molecular mechanisms.Int J Cardiol Heart Vasc. 2016 Mar;10:17-24. doi: 10.1016/j.ijcha.2015.11.004. Int J Cardiol Heart Vasc. 2016. PMID: 27213178 Free PMC article.
-
Apigenin induces DNA damage through the PKCδ-dependent activation of ATM and H2AX causing down-regulation of genes involved in cell cycle control and DNA repair.Biochem Pharmacol. 2012 Dec 15;84(12):1571-80. doi: 10.1016/j.bcp.2012.09.005. Epub 2012 Sep 14. Biochem Pharmacol. 2012. PMID: 22985621 Free PMC article.
References
-
- Akimitsu, N., K. Kamura, S. Tone, A. Sakaguchi, A. Kikuchi, H. Hamamoto, and K. Sekimizu. 2003. Induction of apoptosis by depletion of DNA topoisomerase IIα in mammalian cells. Biochem. Biophys. Res. Commun. 307:301-307. - PubMed
-
- Bharti, A., S. K. Kraeft, M. Gounder, P. Pandey, S. Jin, Z. M. Yuan, S. P. Lees-Miller, R. Weichselbaum, D. Weaver, L. B. Chen, D. Kufe, and S. Kharbanda. 1998. Inactivation of DNA-dependent protein kinase by protein kinase Cδ: implications for apoptosis. Mol. Cell. Biol. 18:6719-6728. - PMC - PubMed
-
- Castedo, M., J. L. Perfettini, T. Roumier, K. Andreau, R. Medema, and G. Kroemer. 2004. Cell death by mitotic catastrophe: a molecular definition. Oncogene 23:2825-2837. - PubMed
-
- Chen, A. Y., and L. F. Liu. 1994. DNA topoisomerases: essential enzymes and lethal targets. Annu. Rev. Pharmacol. Toxicol. 34:191-218. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases