

The evolutionary outcome of sexual conflict
- PMID: 16612889
- PMCID: PMC1569608
- DOI: 10.1098/rstb.2005.1795
The evolutionary outcome of sexual conflict
Abstract
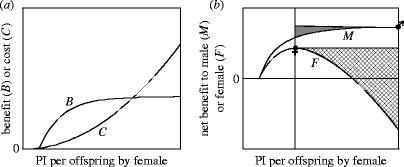
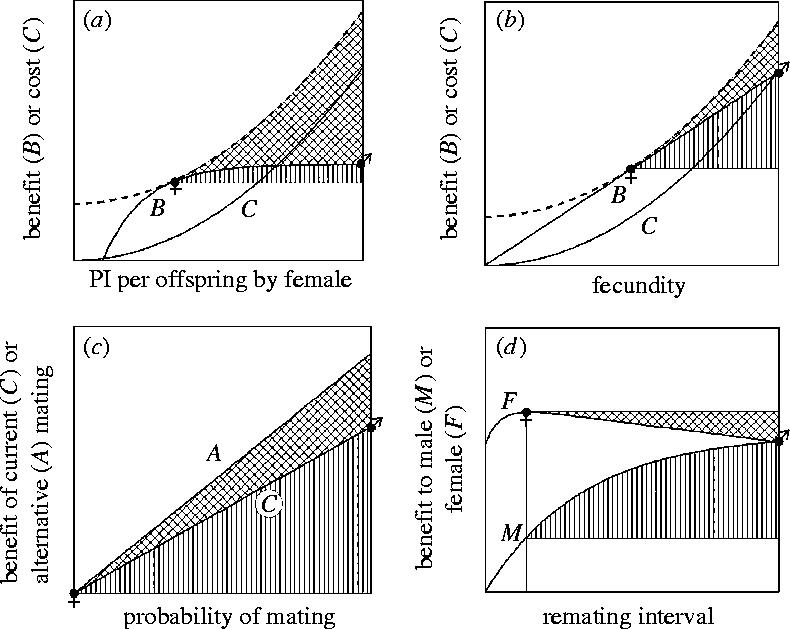
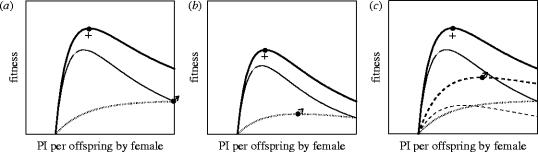
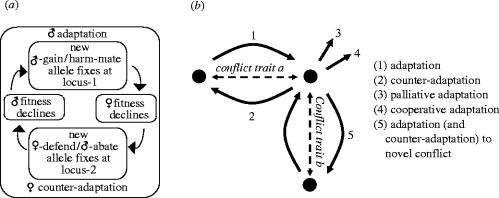
Inter-locus sexual conflict occurs by definition when there is sexually antagonistic selection on a trait so that the optimal trait value differs between the sexes. As a result, there is selection on each sex to manipulate the trait towards its own optimum and resist such manipulation by the other sex. Sexual conflict often leads additionally to the evolution of harmful behaviour and to self-reinforcing and even perpetual sexually antagonistic coevolution. In an attempt to understand the determinants of these different outcomes, I compare two groups of traits-those related to parental investment (PI) and to mating-over which there is sexual conflict, but which have to date been explored by largely separate research traditions. A brief review suggests that sexual conflict over PI, particularly over PI per offspring, leads less frequently to the evolution of manipulative behaviour, and rarely to the evolution of harmful behaviour or to the rapid evolutionary changes which may be symptomatic of sexually antagonistic coevolution. The chief determinants of the evolutionary outcome of sexual conflict are the benefits of manipulation and resistance, the costs of manipulation and resistance, and the feasibility of manipulation. All three of these appear to contribute to the differences in the evolutionary outcome of conflicts over PI and mating. A detailed dissection of the evolutionary changes following from sexual conflict exposes greater complexity than a simple adaptation-counter-adaptation cycle and clarifies the role of harm. Not all of the evolutionary changes that follow from sexual conflict are sexually antagonistic, and harm is not necessary for sexually antagonistic coevolution to occur. In particular, whereas selection on the trait over which there is conflict is by definition sexually antagonistic, collateral harm is usually in the interest of neither sex. This creates the opportunity for palliative adaptations which reduce collateral harm. Failure to recognize that such adaptations are in the interest of both sexes can hinder our understanding of the evolutionary outcome of sexual conflict.
Figures
References
-
- Alatalo R.V, Carlson A, Lundberg A, Ulfstrand S. The conflict between male polygamy and female monogamy: the case of the pied flycatcher Ficedula hypoleuca. Am. Nat. 1981;117:738–753. - DOI
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Research Materials
Miscellaneous