

The dynamics of two- and three-way sexual conflicts over mating
- PMID: 16612892
- PMCID: PMC1569609
- DOI: 10.1098/rstb.2005.1792
The dynamics of two- and three-way sexual conflicts over mating
Abstract
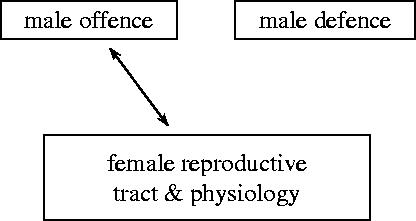
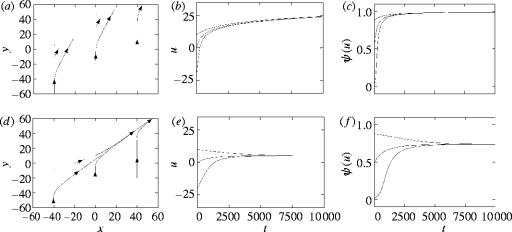
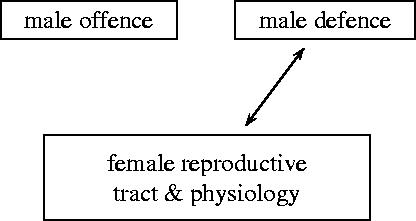
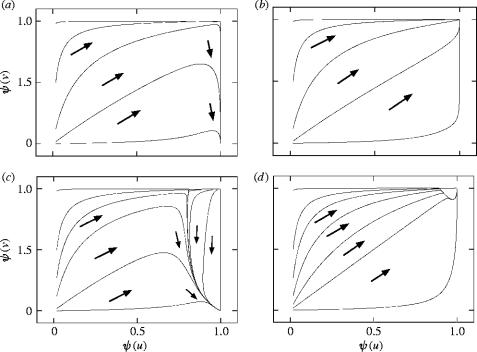
We consider mathematical models describing the evolutionary consequences of antagonistic interactions between male offence, male defence and female reproductive tract and physiology in controlling female mating rate. Overall, the models support previous verbal arguments about the possibility of continuous coevolutionary chase between the sexes driven by two-way (e.g. between male offence and female traits) and three-way (e.g. between male offence, male defence and female traits) inter-sexual antagonistic interactions. At the same time, the models clarify these arguments by identifying various additional potential evolutionary dynamics and important parameters (e.g. genetic variances, female optimum mating rates, strength of selection in females and the relative contributions of first and second males into offspring) and emphasizing the importance of initial conditions. Models also show that sexual conflict can result in the evolution of monandry in an initially polyandrous species and in the evolution of random mating in a population initially exhibiting non-random mating.
Figures




References
-
- Abrams P.A, Harada Y, Matsuda H. On the relationship between quantitative genetic and ESS models. Evolution. 1993;47:982–985. - PubMed
-
- Arnqvist G, Nilsson T. The evolution of polyandry: multiple mating and female fitness in insects. Anim. Behav. 2000;60:145–164. doi:10.1006/anbe.2000.1446 - DOI - PubMed
-
- Arnqvist G, Rowe L. Sexual conflict. Princeton University Press; Princeton, NJ: 2005.
-
- Ball M.A, Parker G.A. Sperm competition games: sperm selection by females. J. Theor. Biol. 2003;224:27–42. doi:10.1016/S0022-5193(03)00118-8 - DOI - PubMed
-
- Chapman T, Arnqvist G, Bangham J, Rowe L. Sexual conflict. Trends Ecol. Evol. 2003;18:41–47. doi:10.1016/S0169-5347(02)00004-6 - DOI
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources