

Sensory exploitation and sexual conflict
- PMID: 16612895
- PMCID: PMC1569614
- DOI: 10.1098/rstb.2005.1790
Sensory exploitation and sexual conflict
Abstract
Much of the literature on male-female coevolution concerns the processes by which male traits and female preferences for these can coevolve and be maintained by selection. There has been less explicit focus on the origin of male traits and female preferences. Here, I argue that it is important to distinguish origin from subsequent coevolution and that insights into the origin can help us appreciate the relative roles of various coevolutionary processes for the evolution of diversity in sexual dimorphism. I delineate four distinct scenarios for the origin of male traits and female preferences that build on past contributions, two of which are based on pre-existing variation in quality indicators among males and two on exploitation of pre-existing sensory biases among females. Recent empirical research, and theoretical models, suggest that origin by sensory exploitation has been widespread. I argue that this points to a key, but perhaps transient, role for sexually antagonistic coevolution (SAC) in the subsequent evolutionary elaboration of sexual traits, because (i) sensory exploitation is often likely to be initially costly for individuals of the exploited sex and (ii) the subsequent evolution of resistance to sensory exploitation should often be associated with costs due to selective constraints. A review of a few case studies is used to illustrate these points. Empirical data directly relevant to the costs of being sensory exploited and the costs of evolving resistance is largely lacking, and I stress that such data would help determining the general importance of sexual conflict and SAC for the evolution of sexual dimorphism.
Figures
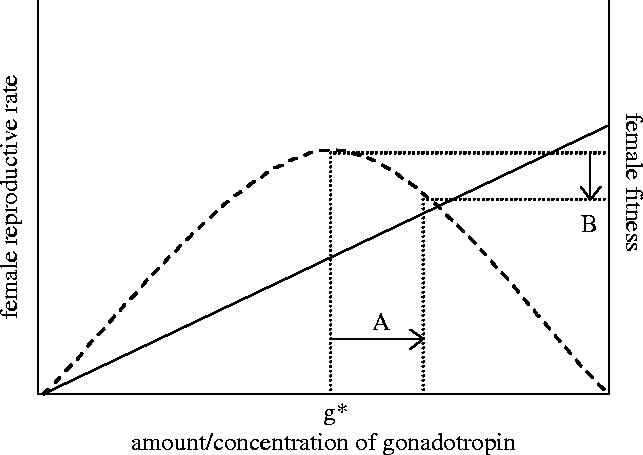
References
-
- Andersson M.Sexual selection1994Princeton University Press; Princeton, NJ
-
- Arak A, Enquist M. Hidden preferences and the evolution of signals. Phil. Trans. R. Soc. B. 1993;340:207–213.
-
- Arnqvist G. Comparative evidence for the evolution of genitalia by sexual selection. Nature. 1998;393:784–786. - DOI
-
- Arnqvist G, Rowe L.Sexual conflict2005Princeton University Press; Princeton, NJ
Publication types
MeSH terms
LinkOut - more resources
Full Text Sources
Miscellaneous