

Extrusion of Ca2+ from mouse motor terminal mitochondria via a Na+-Ca2+ exchanger increases post-tetanic evoked release
- PMID: 16613870
- PMCID: PMC1817729
- DOI: 10.1113/jphysiol.2006.110841
Extrusion of Ca2+ from mouse motor terminal mitochondria via a Na+-Ca2+ exchanger increases post-tetanic evoked release
Abstract
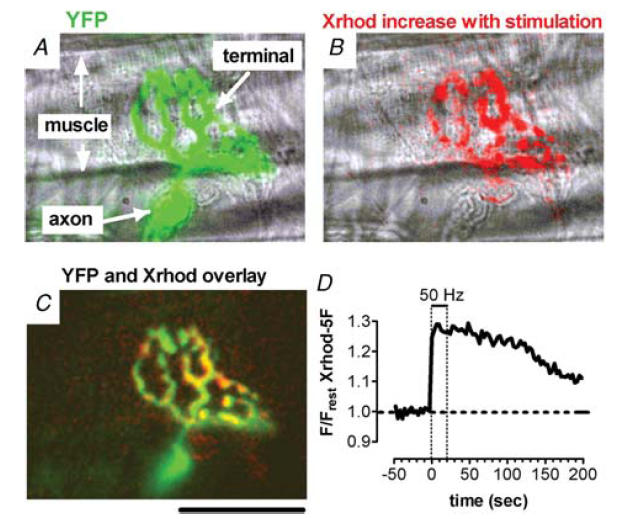
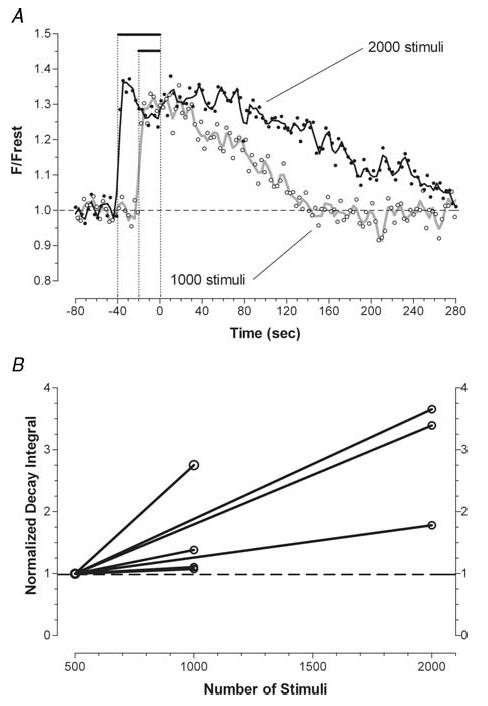
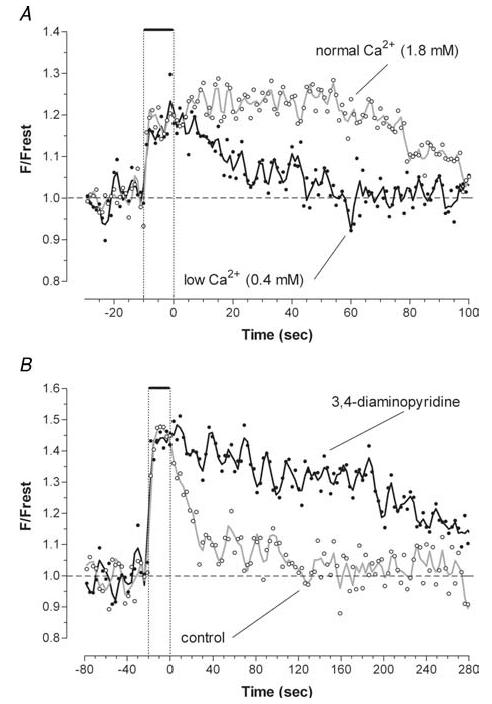
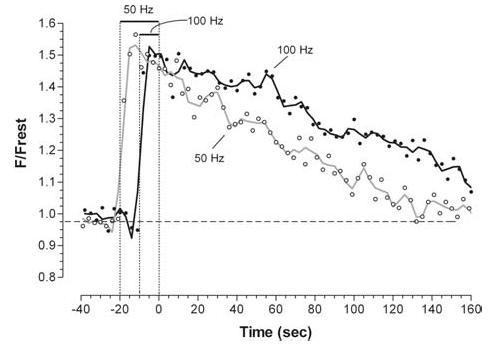
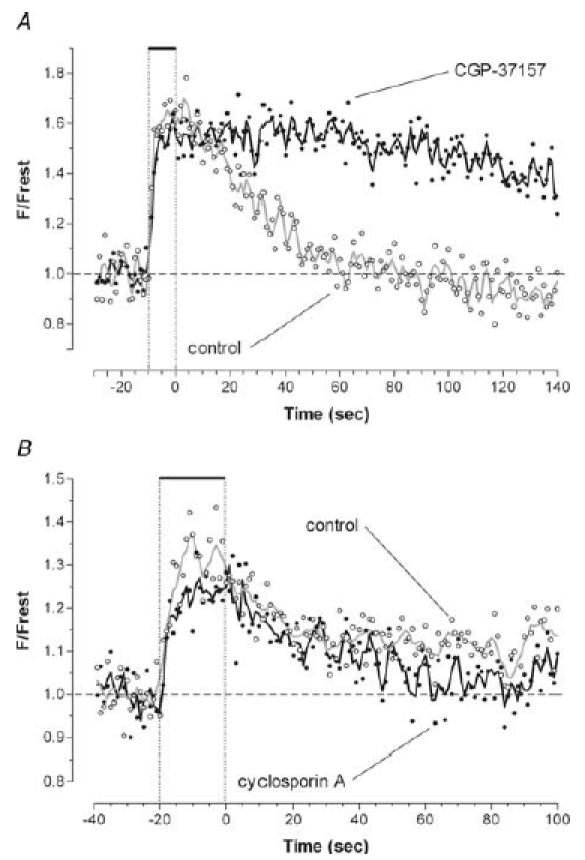
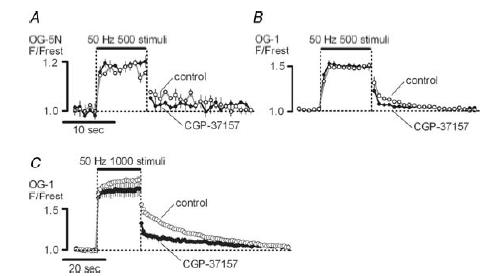
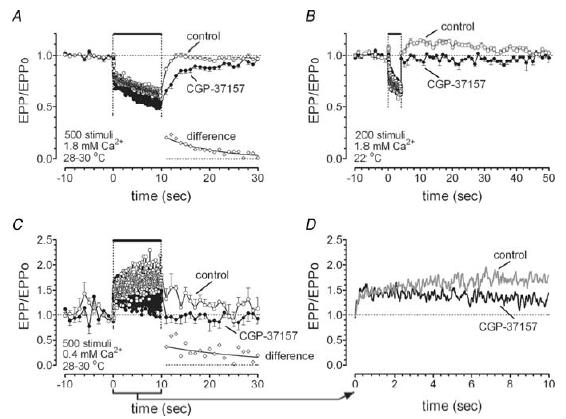
Mitochondria sequester much of the Ca2+ that enters motor nerve terminals during repetitive stimulation at frequencies exceeding 10-20 Hz. We studied the post-stimulation extrusion of Ca2+ from mitochondria by measuring changes in matrix [Ca2+] with fluorescent indicators loaded into motor terminal mitochondria in the mouse levator auris longus muscle. Trains of action potentials at 50 Hz produced a rapid increase in mitochondrial [Ca2+] followed by a plateau, which was usually maintained after the end of the stimulus train and then slowly decayed back to baseline. Increasing the Ca2+ load delivered to the terminal by increasing the number of stimuli (from 500 to 2000) or the stimulation frequency (from 50 to 100 Hz), by increasing bath [Ca2+], or by prolonging the action potential with 3,4-diaminopyridine (100 microM) prolonged the post-stimulation decay of mitochondrial [Ca2+] without increasing the amplitude of the plateau during stimulation. Inhibiting the opening of the mitochondrial permeability transition pore with cyclosporin A (5 microM) had no significant effect on the decay of mitochondrial [Ca2+]. Inhibition of the mitochondrial Na+-Ca2+ exchanger with CGP-37157 (50 microM) dramatically prolonged the post-stimulation decay of mitochondrial [Ca2+], reduced post-stimulation residual cytosolic [Ca2+], and reduced the amplitude of endplate potentials evoked after the end of a stimulus train in the presence of both low and normal bath [Ca2+]. These findings suggest that Ca2+ extrusion from motor terminal mitochondria occurs primarily via the mitochondrial Na+-Ca2+ exchanger and helps to sustain post-tetanic transmitter release at mouse neuromuscular junctions.
Figures
References
-
- Angaut-Petit D, Molgo J, Connold AL, Faille L. The levator auris longus muscle of the mouse: a convenient preparation for studies of short- and long-term presynaptic effects of drugs or toxins. Neurosci Lett. 1987;82:83–88. - PubMed
-
- Argaw A, Desaulniers P, Gardiner PF. Enhanced neuromuscular transmission efficacy in overloaded rat plantaris muscle. Muscle Nerve. 2004;29:97–103. - PubMed
-
- Baron KT, Thayer SA. CGP 37157 modulates mitochondrial Ca2+ homeostasis in cultured rat dorsal root ganglion neurons. Eur J Pharmacol. 1997;340:295–300. - PubMed
-
- Bernardi P. Mitochondrial transport of cations: channels, exchangers, and permeability transition. Physiol Rev. 1999;79:1127–1155. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous