

Activity-dependent synaptic plasticity in the supraoptic nucleus of the rat hypothalamus
- PMID: 16613872
- PMCID: PMC1779752
- DOI: 10.1113/jphysiol.2006.109447
Activity-dependent synaptic plasticity in the supraoptic nucleus of the rat hypothalamus
Abstract
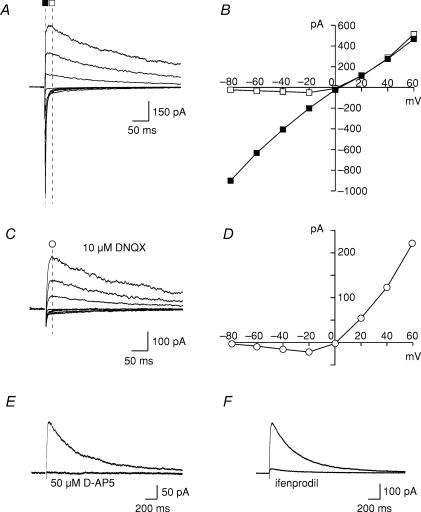
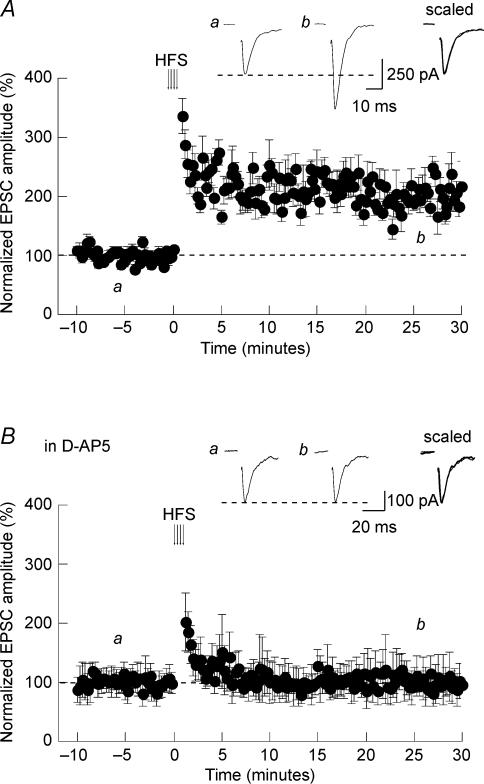
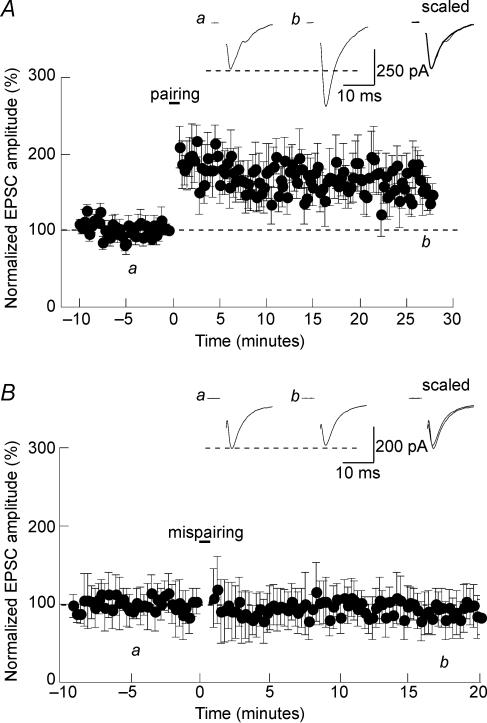
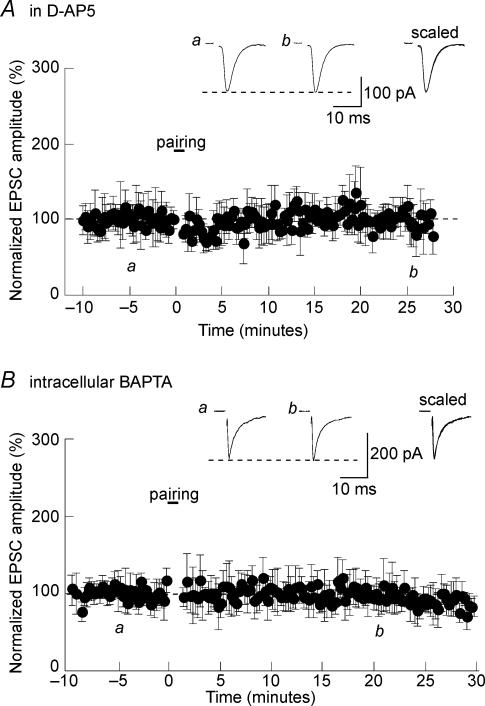
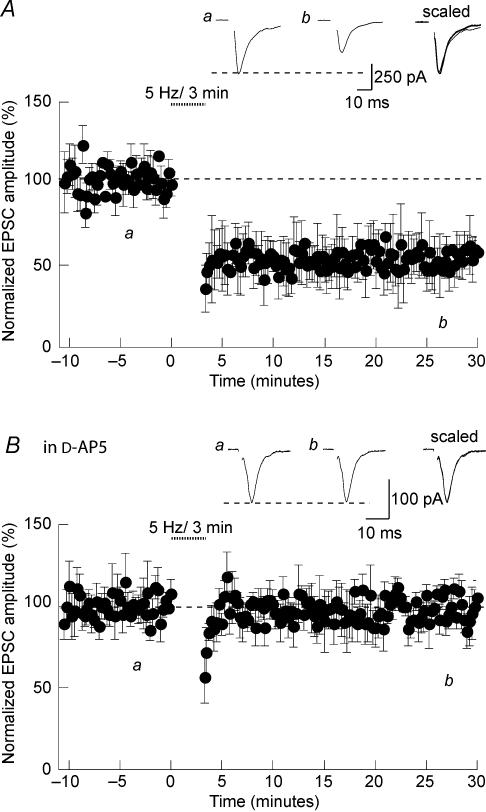
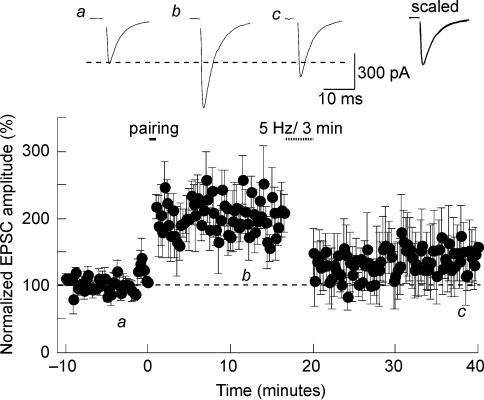
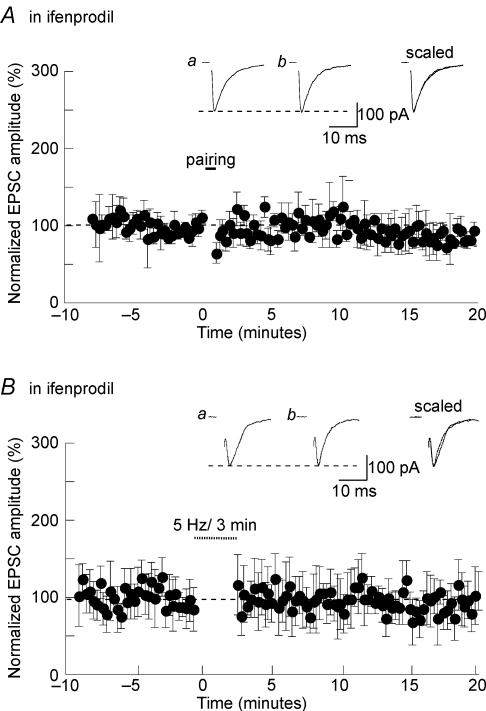
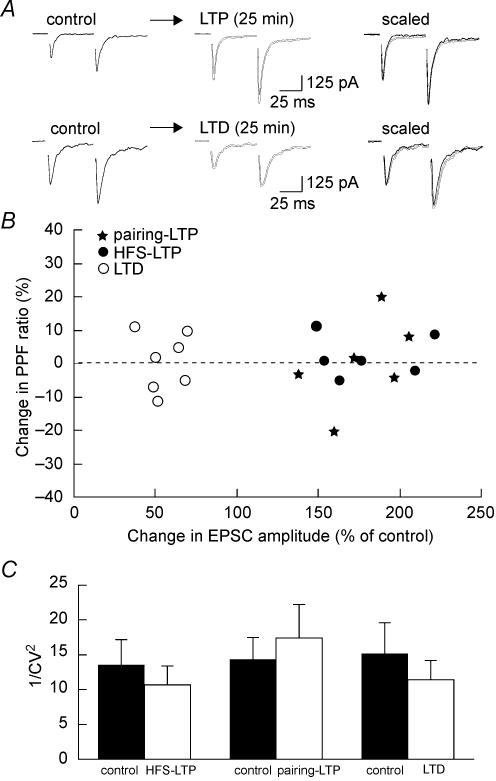
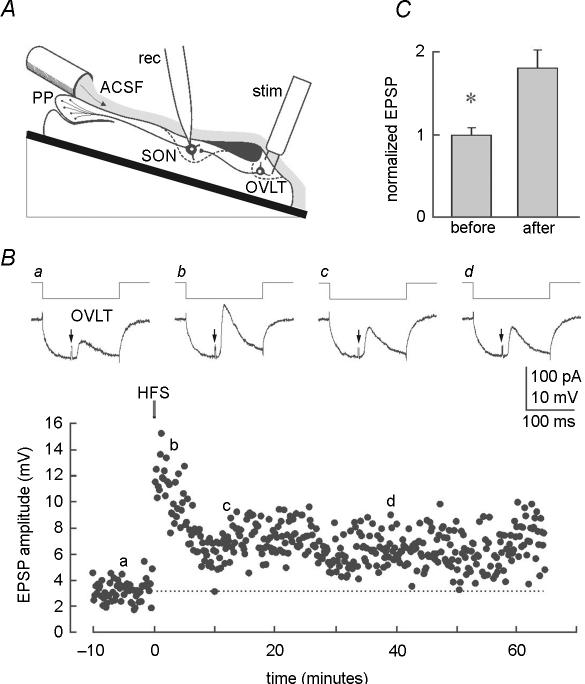
Activity-dependent long-term synaptic changes were investigated at glutamatergic synapses in the supraoptic nucleus (SON) of the rat hypothalamus. In acute hypothalamic slices, high frequency stimulation (HFS) of afferent fibres caused long-term potentiation (LTP) of the amplitude of AMPA receptor-mediated excitatory postsynaptic currents (EPSCs) recorded with the whole-cell patch-clamp technique. LTP was also obtained in response to membrane depolarization paired with mild afferent stimulation. On the other hand, stimulating the inputs at 5 Hz for 3 min at resting membrane potential caused long-term depression (LTD) of excitatory transmission in the SON. These forms of synaptic plasticity required the activation of NMDA receptors since they were abolished in the presence of D-AP5 or ifenprodil, two selective blockers of these receptors. Analysis of paired-pulse facilitation and trial-to-trial variability indicated that LTP and LTD were not associated with changes in the probability of transmitter release, thereby suggesting that the locus of expression of these phenomena was postsynaptic. Using sharp microelectrode recordings in a hypothalamic explant preparation, we found that HFS also generates LTP at functionally defined glutamatergic synapses formed between the organum vasculosum lamina terminalis and SON neurons. Taken together, our findings indicate that glutamatergic synapses in the SON exhibit activity-dependent long-term synaptic changes similar to those prevailing in other brain areas. Such forms of plasticity could play an important role in the context of physiological responses, like dehydration or lactation, where the activity of presynaptic glutamatergic neurons is strongly increased.
Figures
References
-
- Csaki A, Kocsis K, Kiss J, Halasz B. Localization of putative glutamatergic/aspartatergic neurons projecting to the supraoptic nucleus area of the rat hypothalamus. Eur J Neurosci. 2002;16:55–68. - PubMed
-
- Cummings JA, Mulkey RM, Nicoll RA, Malenka RC. Ca2+ signaling requirements for long-term depression in the hippocampus. Neuron. 1996;16:825–833. - PubMed
-
- Gentles SJ. Montreal, Canada: McGill University; 1987. Patterned afferent activity and synaptic plasticity in the magnocellular neurosecretory system. MSc Thesis.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources