

Defocused orientation and position imaging (DOPI) of myosin V
- PMID: 16614073
- PMCID: PMC1458912
- DOI: 10.1073/pnas.0507134103
Defocused orientation and position imaging (DOPI) of myosin V
Abstract
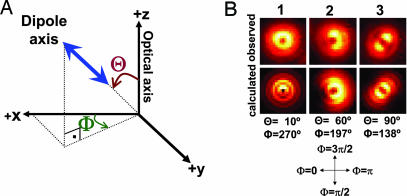
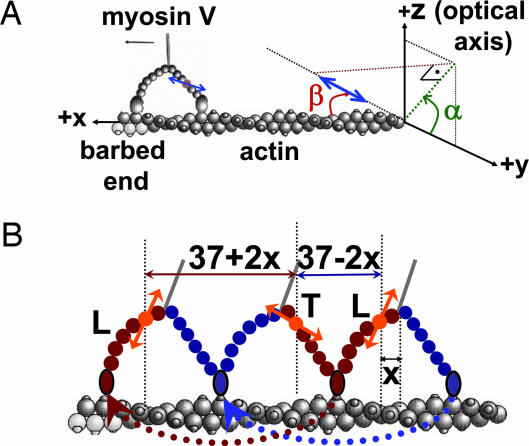
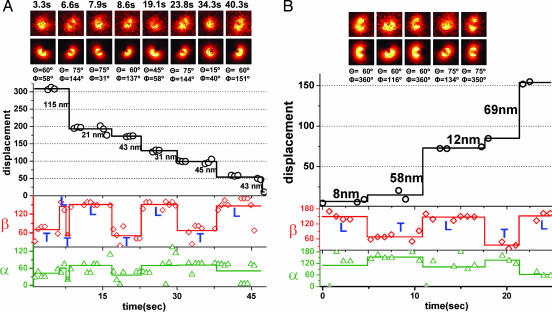
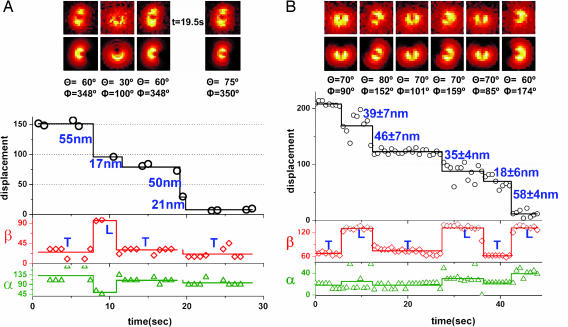
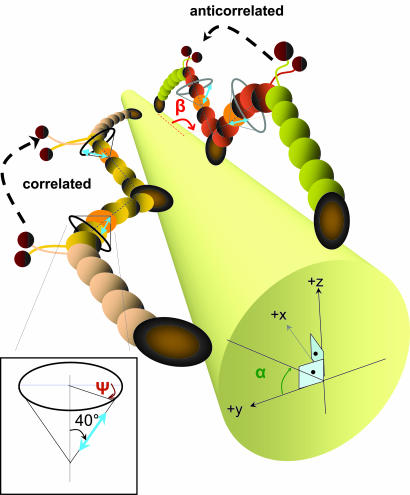
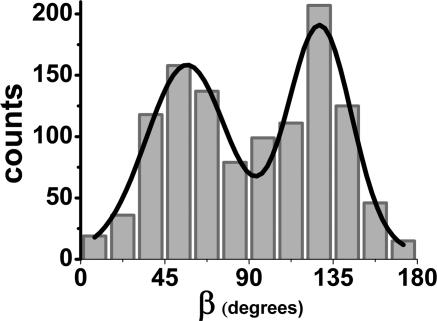
The centroid of a fluorophore can be determined within approximately 1.5-nm accuracy from its focused image through fluorescence imaging with one-nanometer accuracy (FIONA). If, instead, the sample is moved away from the focus, the point-spread-function depends on both the position and 3D orientation of the fluorophore, which can be calculated by defocused orientation and position imaging (DOPI). DOPI does not always yield position accurately, but it is possible to switch back and forth between focused and defocused imaging, thereby getting the centroid and the orientation with precision. We have measured the 3D orientation and stepping behavior of single bifunctional rhodamine probes attached to one of the calmodulins of the light-chain domain (LCD) of myosin V as myosin V moves along actin. Concomitant with large and small steps, the LCD rotates and then dwells in the leading and trailing position, respectively. The probe angle relative to the barbed end of the actin (beta) averaged 128 degrees while the LCD was in the leading state and 57 degrees in the trailing state. The angular difference of 71 degrees represents rotation of LCD around the bound motor domain and is consistent with a 37-nm forward step size of myosin V. When beta changes, the probe rotates +/-27 degrees azimuthally around actin and then rotates back again on the next step. Our results remove degeneracy in angles and the appearance of nontilting lever arms that were reported.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Yildiz A., Forkey J. N., McKinney S. A., Ha T., Goldman Y. E., Selvin P. R. Science. 2003;300:2061–2065. - PubMed
-
- Yildiz A., Tomishige M., Vale R. D., Selvin P. R. Science. 2004;303:676–678. - PubMed
-
- Kural C., Kim H., Syed S., Goshima G., Gelfand V. I., Selvin P. R. Science. 2005;308:1469–1472. - PubMed
-
- Forkey J. N., Quinlan M. E., Shaw M. A., Corrie J. E., Goldman Y. E. Nature. 2003;422:399–404. - PubMed
-
- Bartko A. P., Dickson R. M. J. Phys. Chem. B. 1999;103:3053–3056.
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
