

The continuing challenges of leprosy
- PMID: 16614253
- PMCID: PMC1471987
- DOI: 10.1128/CMR.19.2.338-381.2006
The continuing challenges of leprosy
Abstract
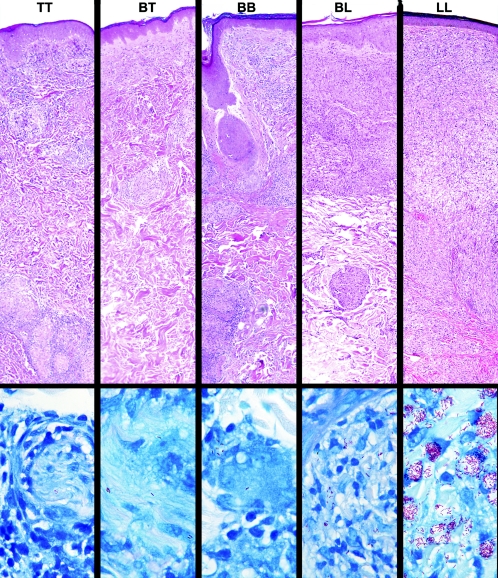
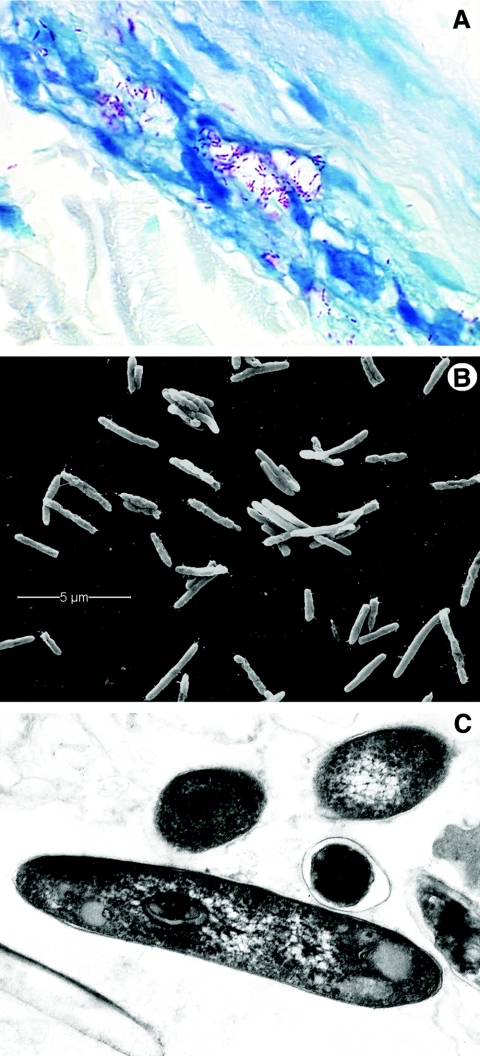
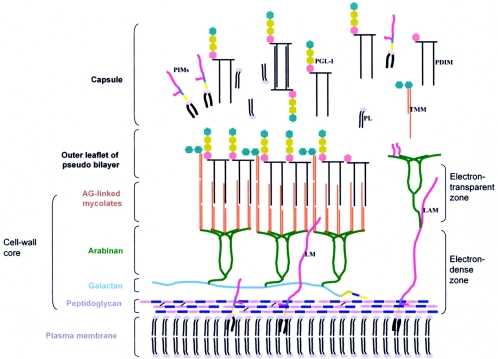
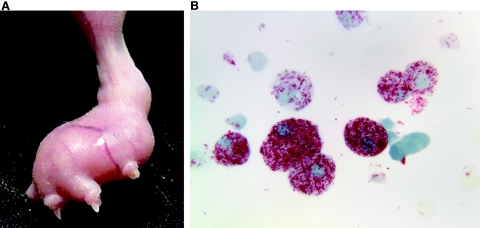
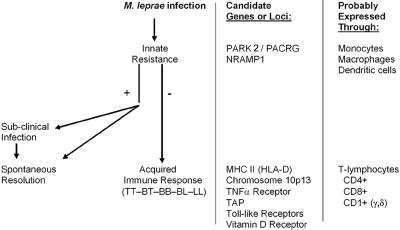
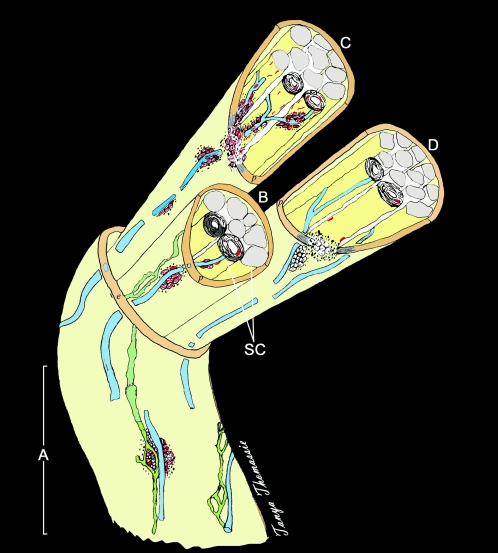
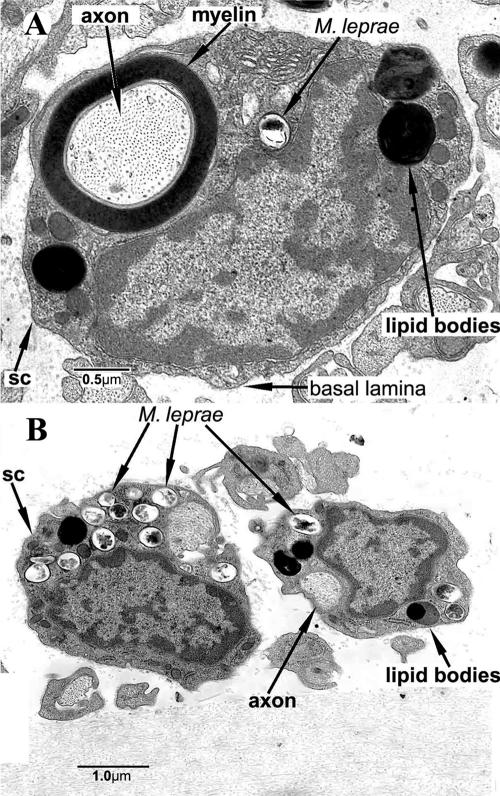
Leprosy is best understood as two conjoined diseases. The first is a chronic mycobacterial infection that elicits an extraordinary range of cellular immune responses in humans. The second is a peripheral neuropathy that is initiated by the infection and the accompanying immunological events. The infection is curable but not preventable, and leprosy remains a major global health problem, especially in the developing world, publicity to the contrary notwithstanding. Mycobacterium leprae remains noncultivable, and for over a century leprosy has presented major challenges in the fields of microbiology, pathology, immunology, and genetics; it continues to do so today. This review focuses on recent advances in our understanding of M. leprae and the host response to it, especially concerning molecular identification of M. leprae, knowledge of its genome, transcriptome, and proteome, its mechanisms of microbial resistance, and recognition of strains by variable-number tandem repeat analysis. Advances in experimental models include studies in gene knockout mice and the development of molecular techniques to explore the armadillo model. In clinical studies, notable progress has been made concerning the immunology and immunopathology of leprosy, the genetics of human resistance, mechanisms of nerve injury, and chemotherapy. In nearly all of these areas, however, leprosy remains poorly understood compared to other major bacterial diseases.
Figures
References
-
- Abel, L., F. O. Sanchez, J. Oberti, N. V. Thuc, L. V. Hoa, V. D. Lap, E. Skamene, P. H. Lagrange, and E. Schurr. 1998. Susceptibility to leprosy is linked to the human NRAMP1 gene. J. Infect. Dis. 177:133-145. - PubMed
-
- Adams, L. B., M. C. Dinauer, D. E. Morgenstern, and J. L. Krahenbuhl. 1997. Comparison of the roles of reactive oxygen and nitrogen intermediates in the host response to Mycobacterium tuberculosis using transgenic mice. Tubercle Lung Dis. 7 8:237-246. - PubMed
-
- Adams, L. B., S. G. Franzblau, Z. Vavrin, J. B. Hibbs, Jr., and J. L. Krahenbuhl. 1991. l-Arginine-dependent macrophage effector functions inhibit metabolic activity of Mycobacterium leprae. J. Immunol. 147:1642-1646. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical
