

Development and evolution of the vestibular sensory apparatus of the mammalian ear
- PMID: 16614470
- PMCID: PMC3901525
Development and evolution of the vestibular sensory apparatus of the mammalian ear
Abstract
Herein, we will review molecular aspects of vestibular ear development and present them in the context of evolutionary changes and hair cell regeneration. Several genes guide the development of anterior and posterior canals. Although some of these genes are also important for horizontal canal development, this canal strongly depends on a single gene, Otx1. Otx1 also governs the segregation of saccule and utricle. Several genes are essential for otoconia and cupula formation, but protein interactions necessary to form and maintain otoconia or a cupula are not yet understood. Nerve fiber guidance to specific vestibular end-organs is predominantly mediated by diffusible neurotrophic factors that work even in the absence of differentiated hair cells. Neurotrophins, in particular Bdnf, are the most crucial attractive factor released by hair cells. If Bdnf is misexpressed, fibers can be redirected away from hair cells. Hair cell differentiation is mediated by Atoh1. However, Atoh1 may not initiate hair cell precursor formation. Resolving the role of Atoh1 in postmitotic hair cell precursors is crucial for future attempts in hair cell regeneration. Additional analyses are needed before gene therapy can help regenerate hair cells, restore otoconia, and reconnect sensory epithelia to the brain.
Figures
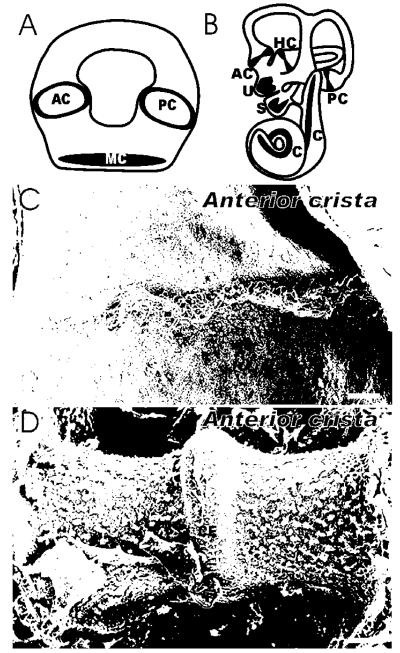





Similar articles
-
Atoh1 null mice show directed afferent fiber growth to undifferentiated ear sensory epithelia followed by incomplete fiber retention.Dev Dyn. 2005 Jun;233(2):570-83. doi: 10.1002/dvdy.20370. Dev Dyn. 2005. PMID: 15844198 Free PMC article.
-
Mutant mice reveal the molecular and cellular basis for specific sensory connections to inner ear epithelia and primary nuclei of the brain.Hear Res. 2005 Aug;206(1-2):52-63. doi: 10.1016/j.heares.2004.11.025. Hear Res. 2005. PMID: 16080998 Free PMC article. Review.
-
Otx1 null mutant mice show partial segregation of sensory epithelia comparable to lamprey ears.Dev Genes Evol. 2001 Sep;211(8-9):388-96. doi: 10.1007/s004270100166. Dev Genes Evol. 2001. PMID: 11685572
-
Analysis of rat vestibular hair cell development and regeneration using calretinin as an early marker.J Neurosci. 1997 Nov 1;17(21):8270-82. doi: 10.1523/JNEUROSCI.17-21-08270.1997. J Neurosci. 1997. PMID: 9334402 Free PMC article.
-
Comparative biology of the amniote vestibular utricle.Hear Res. 2024 Jul;448:109035. doi: 10.1016/j.heares.2024.109035. Epub 2024 May 19. Hear Res. 2024. PMID: 38763033 Review.
Cited by
-
Appropriate Vestibular Stimulation in Children and Adolescents-A Prerequisite for Normal Cognitive, Motor Development and Bodily Homeostasis-A Review.Children (Basel). 2023 Dec 19;11(1):2. doi: 10.3390/children11010002. Children (Basel). 2023. PMID: 38275423 Free PMC article. Review.
-
Modality-specific effects of threat on self-motion perception.BMC Biol. 2024 May 23;22(1):120. doi: 10.1186/s12915-024-01911-3. BMC Biol. 2024. PMID: 38783286 Free PMC article.
-
Pax2-Islet1 Transgenic Mice Are Hyperactive and Have Altered Cerebellar Foliation.Mol Neurobiol. 2017 Mar;54(2):1352-1368. doi: 10.1007/s12035-016-9716-6. Epub 2016 Feb 3. Mol Neurobiol. 2017. PMID: 26843111 Free PMC article.
-
Developmental profiling of spiral ganglion neurons reveals insights into auditory circuit assembly.J Neurosci. 2011 Jul 27;31(30):10903-18. doi: 10.1523/JNEUROSCI.2358-11.2011. J Neurosci. 2011. PMID: 21795542 Free PMC article.
-
Filling the silent void: genetic therapies for hearing impairment.Genet Res Int. 2012;2012:748698. doi: 10.1155/2012/748698. Epub 2012 Dec 4. Genet Res Int. 2012. PMID: 23304527 Free PMC article.
References
-
- Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, Levi-Acobas F, Cruaud C, Le Merrer M, Mathieu M, Konig R, Vigneron J, Weissenbach J, Petit C, Weil D. Clustering of mutations responsible for branchio-oto-renal (BOR) syndrome in the eyes absent homologous region (eyaHR) of EYA1. Hum Mol Genet. 1997;6:2247–2255. - PubMed
-
- Abdelhak S, Kalatzis V, Heilig R, Compain S, Samson D, Vincent C, Weil D, Cruaud C, Sahly I, Leibovici M, Bitner-Glindzicz M, Francis M, Lacombe D, Vigneron J, Charachon R, Boven K, Bedbeder P, Van Regemorter N, Weissenbach J, Petit C. A human homologue of the Drosophila eyes absent gene underlies branchio-oto-renal (BOR) syndrome and identifies a novel gene family. Nat Genet. 1997;15:157–164. - PubMed
-
- Agerman K, Hjerling-Leffler J, Blanchard MP, Scarfone E, Canlon B, Nosrat C, Ernfors P. BDNF gene replacement reveals multiple mechanisms for establishing neurotrophin specificity during sensory nervous system development. Development. 2003;130:1479–1491. - PubMed
-
- Alvarez Y, Alonso MT, Vendrell V, Zelarayan LC, Chamero P, Theil T, Bosl MR, Kato S, Maconochie M, Riethmacher D, Schimmang T. Requirements for FGF3 and FGF10 during inner ear formation. Development. 2003;130:6329–6338. - PubMed
-
- Baker CV, Bronner-Fraser M. Vertebrate cranial placodes I. Embryonic induction. Dev Biol. 2001;232:1–61. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Medical