

Semisynthesis and characterization of mammalian thioredoxin reductase
- PMID: 16618105
- PMCID: PMC2570056
- DOI: 10.1021/bi0517887
Semisynthesis and characterization of mammalian thioredoxin reductase
Abstract
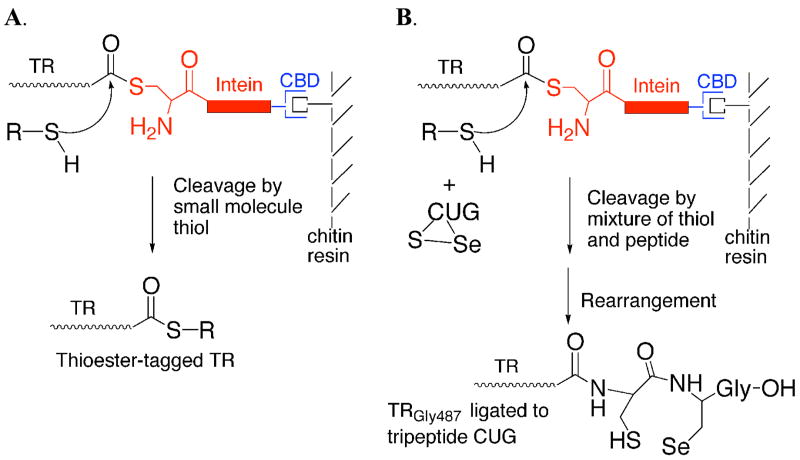
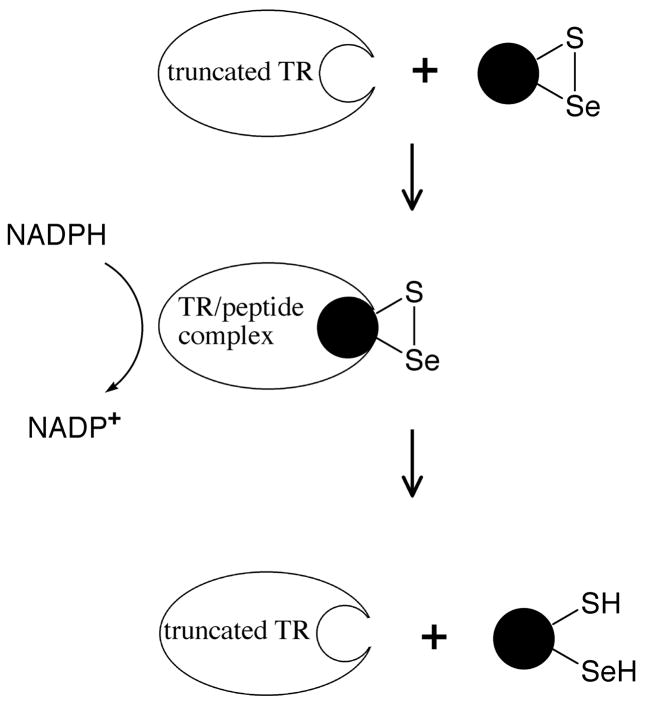
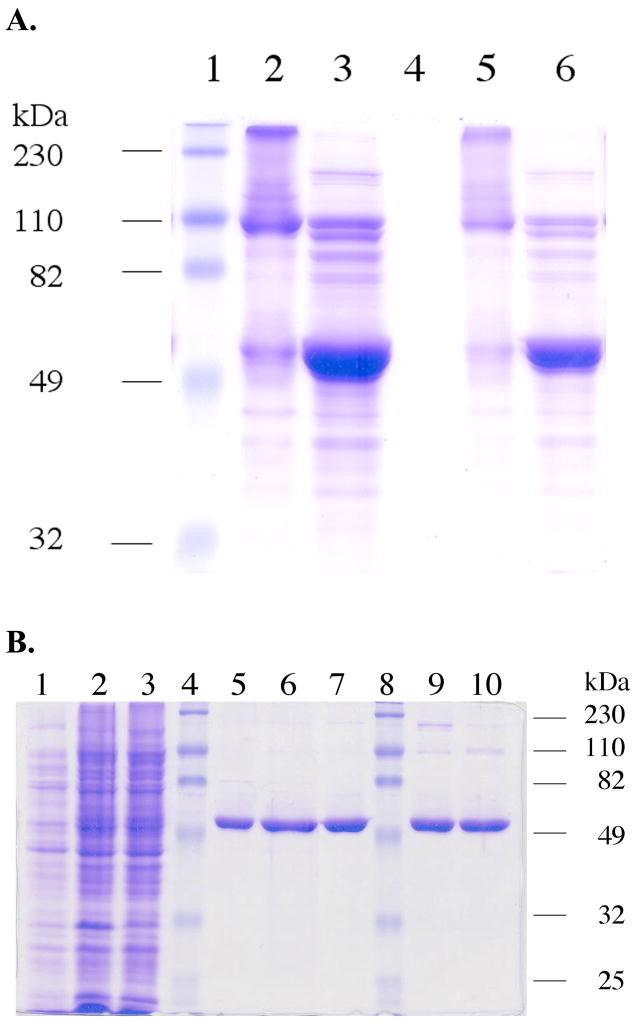
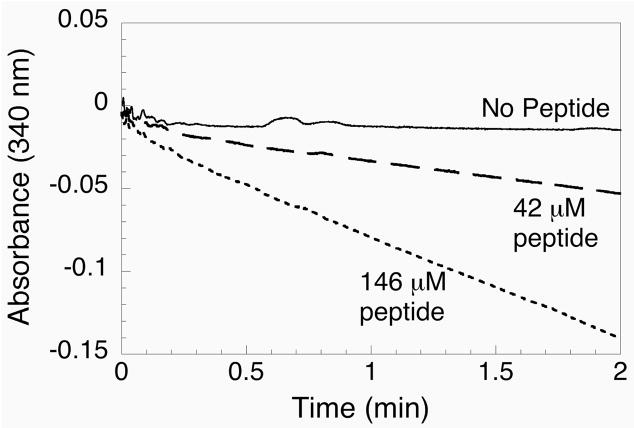
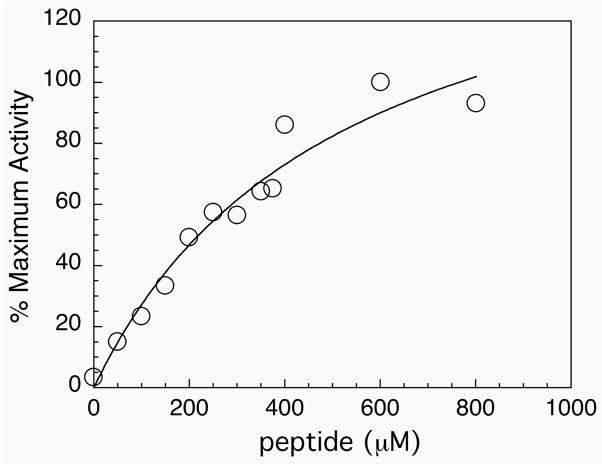
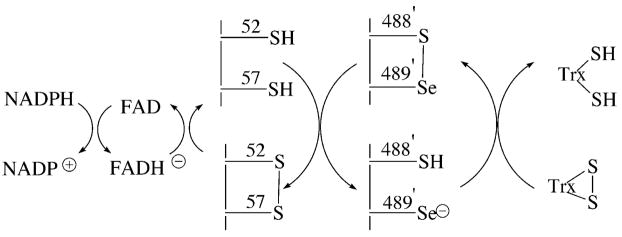
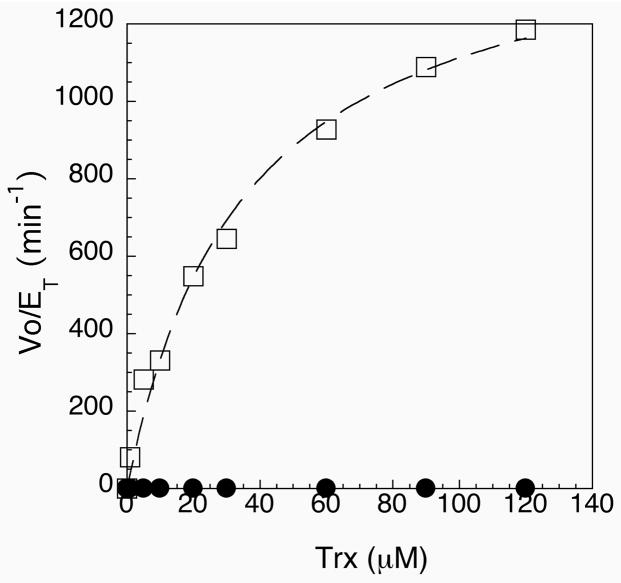
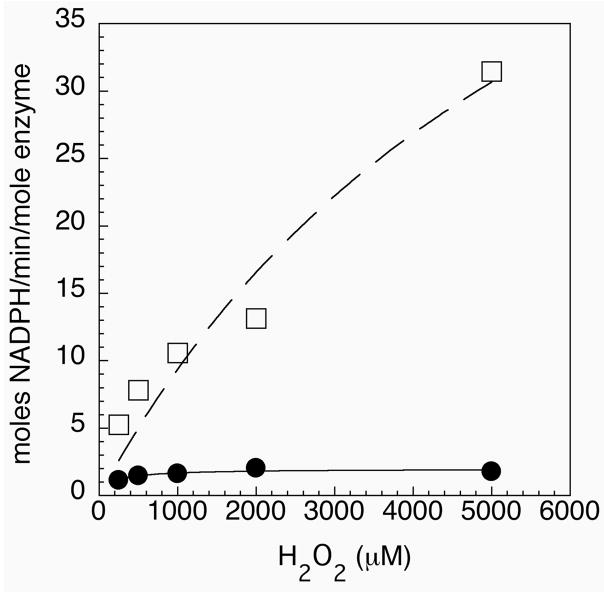
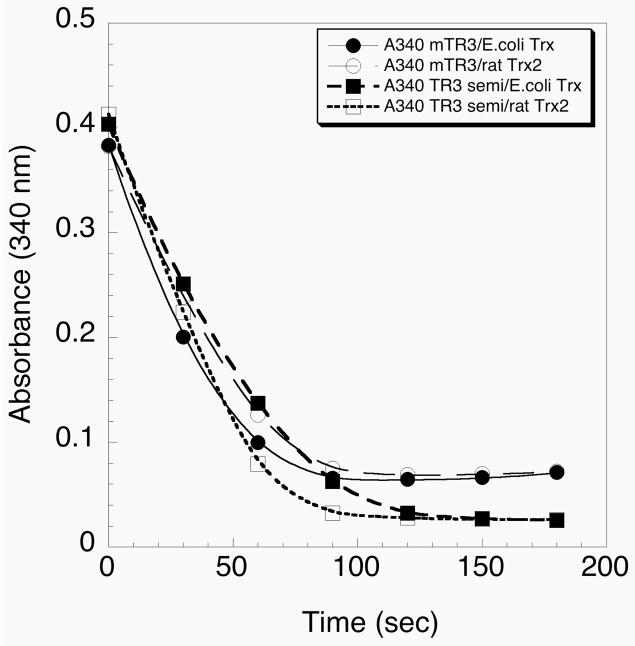
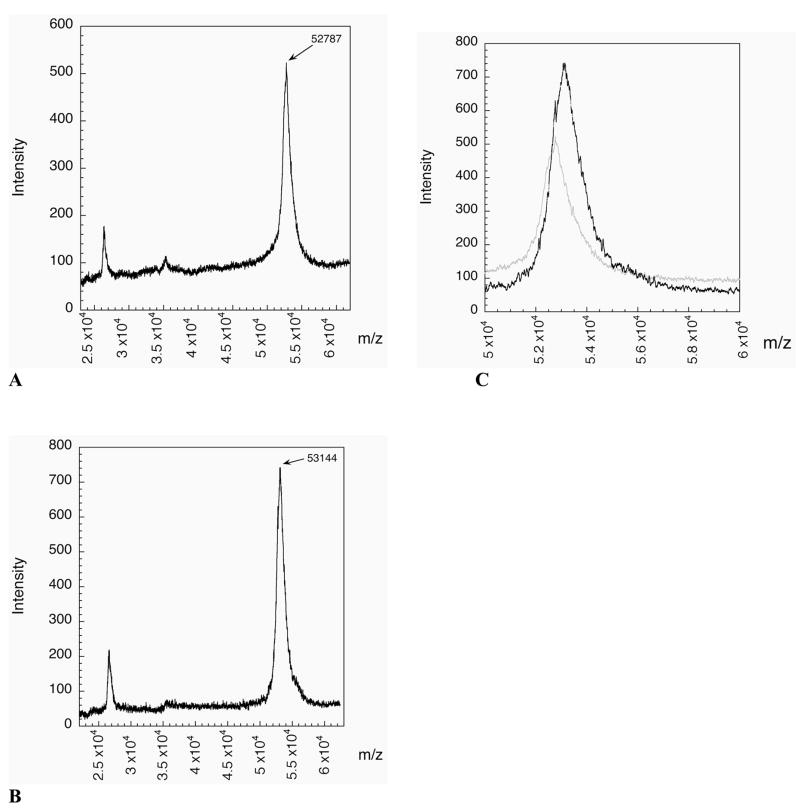
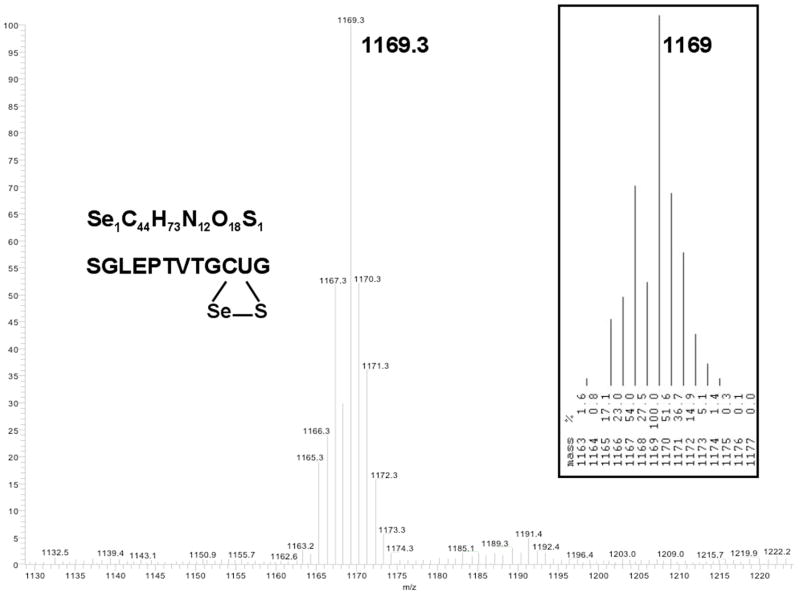
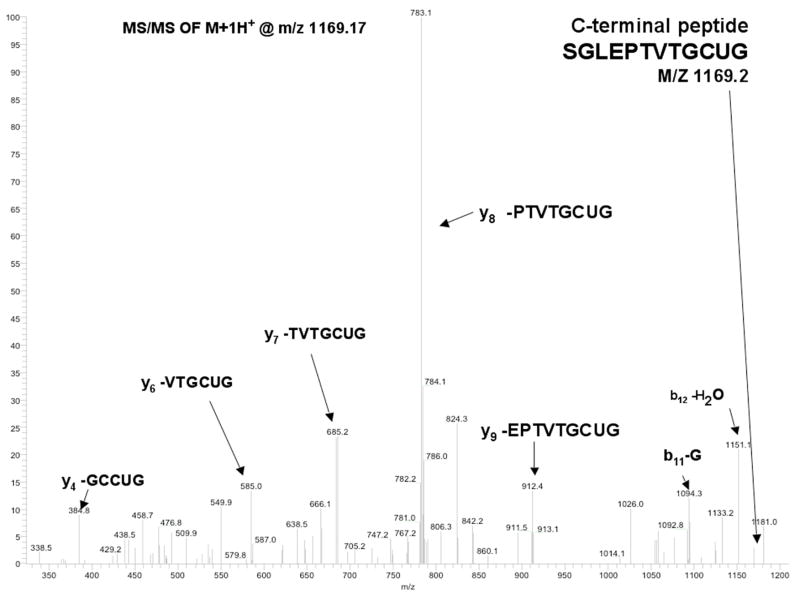
Thioredoxin reductase and thioredoxin constitute the cellular thioredoxin system, which provides reducing equivalents to numerous intracellular target disulfides. Mammalian thioredoxin reductase contains the rare amino acid selenocysteine. Known as the "21st" amino acid, selenocysteine is inserted into proteins by recoding UGA stop codons. Some model eukaryotic organisms lack the ability to insert selenocysteine, and prokaryotes have a recoding apparatus different from that of eukaryotes, thus making heterologous expression of mammalian selenoproteins difficult. Here, we present a semisynthetic method for preparing mammalian thioredoxin reductase. This method produces the first 487 amino acids of mouse thioredoxin reductase-3 as an intein fusion protein in Escherichia coli cells. The missing C-terminal tripeptide containing selenocysteine is then ligated to the thioester-tagged protein by expressed protein ligation. The semisynthetic version of thioredoxin reductase that we produce in this manner has k(cat) values ranging from 1500 to 2220 min(-)(1) toward thioredoxin and has strong peroxidase activity, indicating a functional form of the enzyme. We produced the semisynthetic thioredoxin reductase with a total yield of 24 mg from 6 L of E. coli culture (4 mg/L). This method allows production of a fully functional, semisynthetic selenoenzyme that is amenable to structure-function studies. A second semisynthetic system is also reported that makes use of peptide complementation to produce a partially active enzyme. The results of our peptide complementation studies reveal that a tetrapeptide that cannot ligate to the enzyme (Ac-Gly-Cys-Sec-Gly) can form a noncovalent complex with the truncated enzyme to form a weak complex. This noncovalent peptide-enzyme complex has 350-500-fold lower activity than the semisynthetic enzyme produced by peptide ligation.
Figures
References
-
- Holmgren A. Thioredoxin and glutaredoxin systems. J Biol Chem. 1989;264:13963–13966. - PubMed
-
- Arnér ES, Holmgren A. Physiological functions of thioredoxin and thioredoxin reductase. Eur J Biochem. 2000;267:6102–6109. - PubMed
-
- Atkins JF, Gesteland RF. The twenty-first amino acid. Nature. 2000;407:463–465. - PubMed
-
- Bock A, Forchhammer K, Heider J, Leinfelder W, Sawers G, Veprek B, Zinoni F. Selenocysteine: the 21st amino acid. Mol Microbiol. 1991;5:515–520. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous
