

Feedback control of morphogenesis in fungi by aromatic alcohols
- PMID: 16618799
- PMCID: PMC1472474
- DOI: 10.1101/gad.1411806
Feedback control of morphogenesis in fungi by aromatic alcohols
Abstract
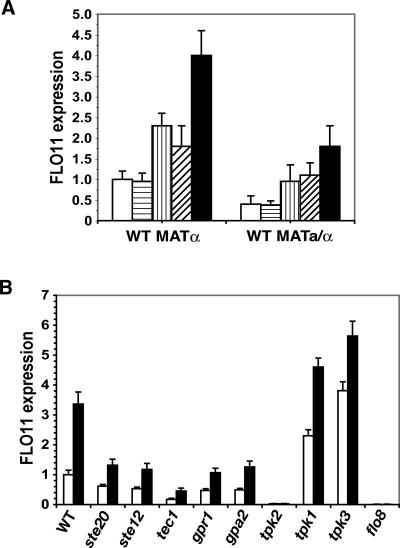
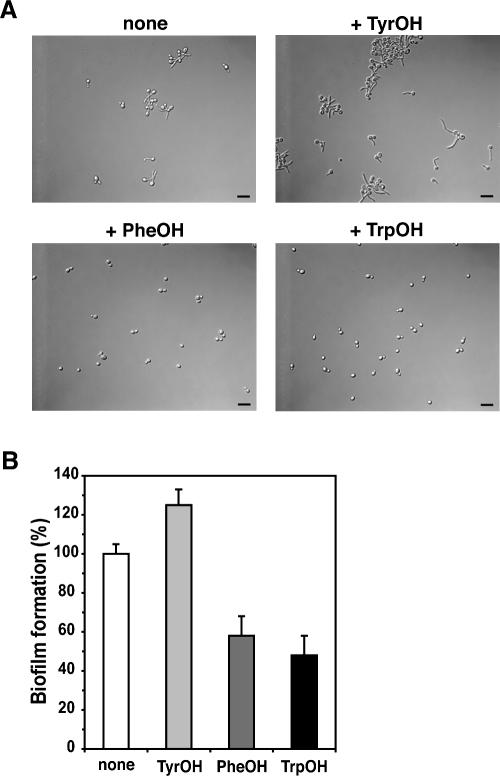
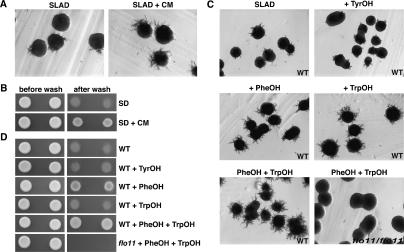
Many fungi undergo a developmental transition from a unicellular yeast form to an invasive filamentous form in response to environmental cues. Here we describe a quorum signaling pathway that links environmental sensing to morphogenesis in Saccharomyces cerevisiae. Saccharomyces cells secrete aromatic alcohols that stimulate morphogenesis by inducing the expression of FLO11 through a Tpk2p-dependent mechanism. Mutants defective in synthesis of these alcohols show reduced filamentous growth, which is partially suppressed by the addition of these aromatic alcohols. The production of these auto signaling alcohols is regulated by nitrogen: High ammonia restricts it by repressing the expression of their biosynthetic pathway, whereas nitrogen-poor conditions activate it. Moreover, the production of these aromatic alcohols is controlled by cell density and subjected to positive feedback regulation, which requires the transcription factor Aro80p. These interactions define a quorum-sensing circuit that allows Saccharomyces to respond to both cell density and the nutritional state of the environment. These same autoregulatory molecules do not evoke the morphological switch in Candida albicans, suggesting that these molecular signals are species-specific.
Figures




Comment in
-
Eukaryotes learn how to count: quorum sensing by yeast.Genes Dev. 2006 May 1;20(9):1045-9. doi: 10.1101/gad.1432906. Genes Dev. 2006. PMID: 16651650 Review. No abstract available.
References
-
- Aubry L., Firtel R. Integration of signaling networks that regulate Dictyostelium differentiation. Annu. Rev. Cell Dev. Biol. 1999;15:469–517. - PubMed
-
- Bassler B.L. Small talk. Cell-to-cell communication in bacteria. Cell. 2002;109:421–424. - PubMed
-
- Biswas K., Morschhauser J. The Mep2p ammonium permease controls nitrogen starvation-induced filamentous growth in Candida albicans. Mol. Microbiol. 2005;56:649–669. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases