

Definition of the bacterial N-glycosylation site consensus sequence
- PMID: 16619027
- PMCID: PMC1456941
- DOI: 10.1038/sj.emboj.7601087
Definition of the bacterial N-glycosylation site consensus sequence
Abstract
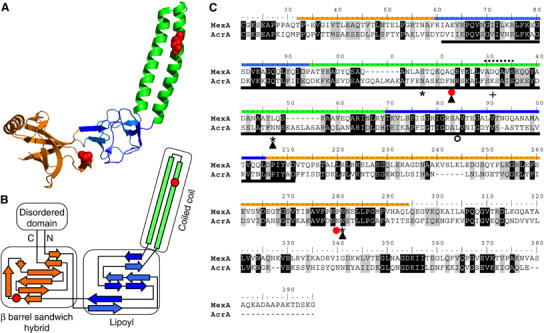
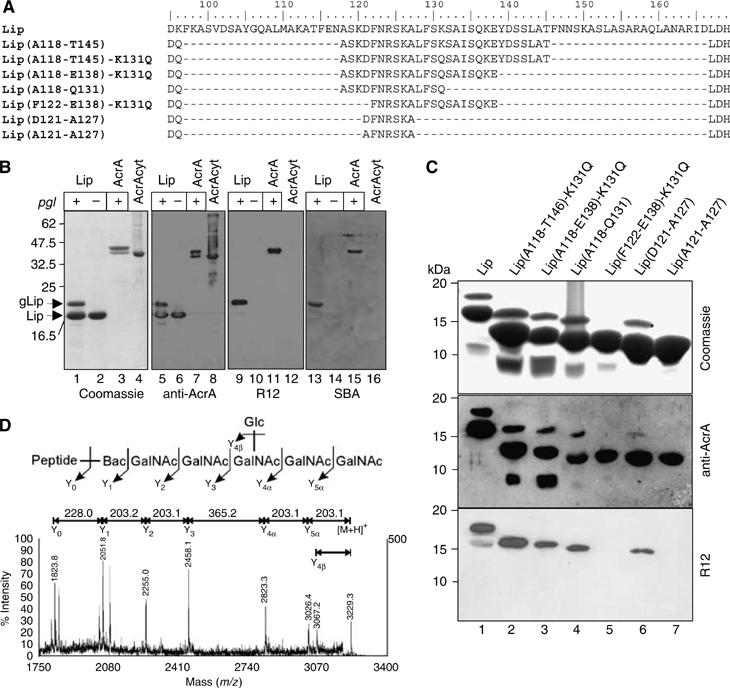
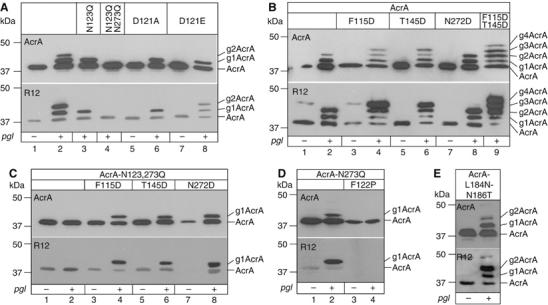
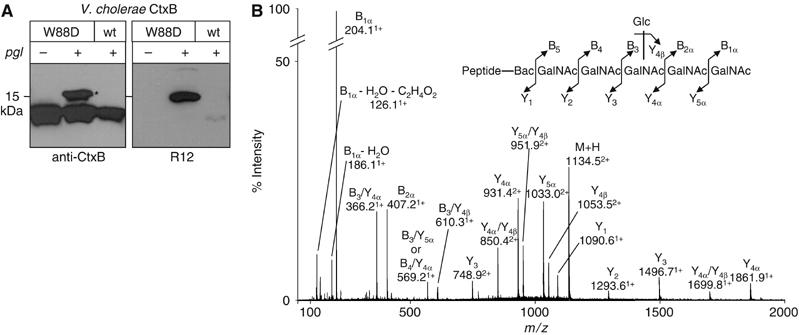
The Campylobacter jejuni pgl locus encodes an N-linked protein glycosylation machinery that can be functionally transferred into Escherichia coli. In this system, we analyzed the elements in the C. jejuni N-glycoprotein AcrA required for accepting an N-glycan. We found that the eukaryotic primary consensus sequence for N-glycosylation is N terminally extended to D/E-Y-N-X-S/T (Y, X not equalP) for recognition by the bacterial oligosaccharyltransferase (OST) PglB. However, not all consensus sequences were N-glycosylated when they were either artificially introduced or when they were present in non-C. jejuni proteins. We were able to produce recombinant glycoproteins with engineered N-glycosylation sites and confirmed the requirement for a negatively charged side chain at position -2 in C. jejuni N-glycoproteins. N-glycosylation of AcrA by the eukaryotic OST in Saccharomyces cerevisiae occurred independent of the acidic residue at the -2 position. Thus, bacterial N-glycosylation site selection is more specific than the eukaryotic equivalent with respect to the polypeptide acceptor sequence.
Figures

References
-
- Akama H, Matsuura T, Kashiwagi S, Yoneyama H, Narita S, Tsukihara T, Nakagawa A, Nakae T (2004) Crystal structure of the membrane fusion protein, MexA, of the multidrug transporter in Pseudomonas aeruginosa. J Biol Chem 279: 25939–25942 - PubMed
-
- Ben-Dor S, Esterman N, Rubin E, Sharon N (2004) Biases and complex patterns in the residues flanking protein N-glycosylation sites. Glycobiology 14: 95–101 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
