

An aging pathway controls the TrkA to p75NTR receptor switch and amyloid beta-peptide generation
- PMID: 16619032
- PMCID: PMC1456930
- DOI: 10.1038/sj.emboj.7601062
An aging pathway controls the TrkA to p75NTR receptor switch and amyloid beta-peptide generation
Abstract
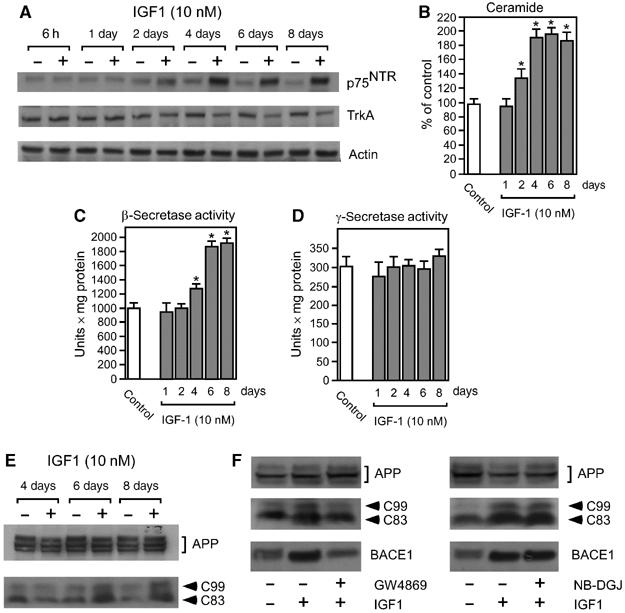
Aging of the brain is characterized by marked changes in the expression levels of the neurotrophin receptors, TrkA and p75(NTR). An expression pattern in which TrkA predominates in younger animals switches to one in which p75(NTR) predominates in older animals. This TrkA-to-p75(NTR) switch is accompanied by activation of the second messenger ceramide, stabilization of beta-site amyloid precursor protein-cleaving enzyme-1 (BACE1), and increased production of amyloid beta-peptide (Abeta). Here, we show that the insulin-like growth factor-1 receptor (IGF1-R), the common regulator of lifespan and age-related events in many different organisms, is responsible for the TrkA-to-p75(NTR) switch in both human neuroblastoma cell lines and primary neurons from mouse brain. The signaling pathway that controls the level of TrkA and p75(NTR) downstream of the IGF1-R requires IRS2, PIP3/Akt, and is under the control of PTEN and p44, the short isoform of p53. We also show that hyperactivation of IGF1-R signaling in p44 transgenic animals, which show an accelerated form of aging, is characterized by early TrkA-to-p75(NTR) switch and increased production of Abeta in the brain.
Figures




References
-
- Bondy CA, Cheng CM (2004) Signaling by insulin-like growth factor 1 in brain. Eur J Pharmacol 490: 25–31 - PubMed
-
- Bowen RL, Verdile G, Liu T, Parlow AF, Perry G, Smith MA, Martins RN, Atwood CS (2004) Luteinizing hormone, a reproductive regulator that modulates the processing of amyloid-beta precursor protein and amyloid-beta deposition. J Biol Chem 279: 20539–20545 - PubMed
-
- Campisi J (2004) Fragile fugue: p53 in aging, cancer and IGF signaling. Nat Med 10: 231–232 - PubMed
-
- Carro E, Torres-Aleman I (2004) The role of insulin and insulin-like growth factor I in the molecular and cellular mechanisms underlying the pathology of Alzheimer's disease. Eur J Pharmacol 490: 127–133 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
Miscellaneous
