

Specific modification of a Na+ binding site in NADH:quinone oxidoreductase from Klebsiella pneumoniae with dicyclohexylcarbodiimide
- PMID: 16621819
- PMCID: PMC1447468
- DOI: 10.1128/JB.188.9.3264-3272.2006
Specific modification of a Na+ binding site in NADH:quinone oxidoreductase from Klebsiella pneumoniae with dicyclohexylcarbodiimide
Abstract
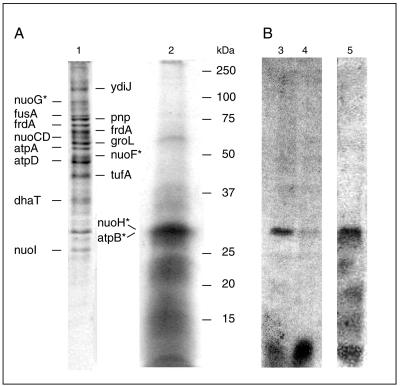
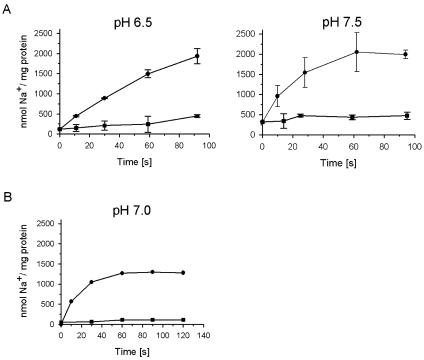
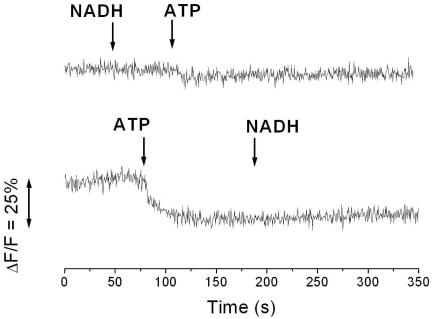
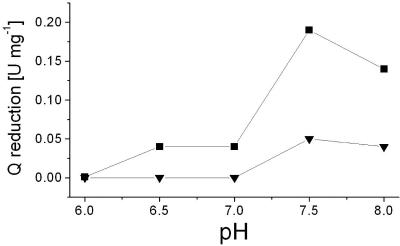
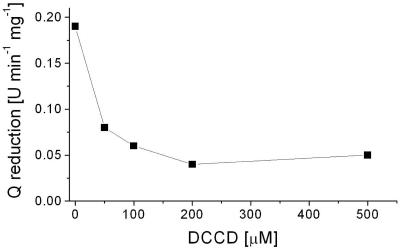
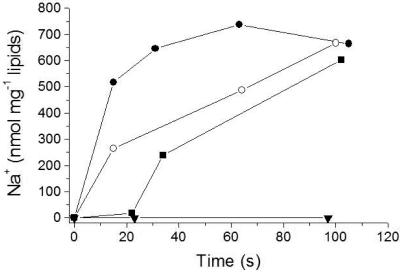
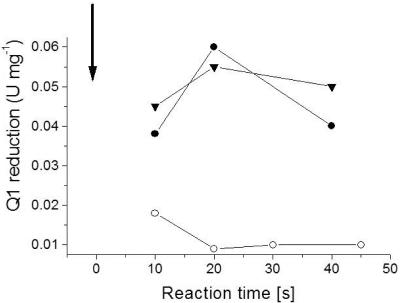
The respiratory NADH:quinone oxidoreductase (complex I) (NDH-1) is a multisubunit enzyme that translocates protons (or in some cases Na+) across energy-conserving membranes from bacteria or mitochondria. We studied the reaction of the Na+-translocating complex I from the enterobacterium Klebsiella pneumoniae with N,N'-dicyclohexylcarbodiimide (DCCD), with the aim of identifying a subunit critical for Na+ binding. At low Na+ concentrations (0.6 mM), DCCD inhibited both quinone reduction and Na+ transport by NDH-1 concurrent with the covalent modification of a 30-kDa polypeptide. In the presence of 50 mM Na+, NDH-1 was protected from inhibition by DCCD, and the modification of the 30-kDa polypeptide with [14C]DCCD was prevented, indicating that Na+ and DCCD competed for the binding to a critical carboxyl group in NDH-1. The 30-kDa polypeptide was assigned to NuoH, the homologue of the ND1 subunit from mitochondrial complex I. It is proposed that Na+ binds to the NuoH subunit during NADH-driven Na+ transport by NDH-1.
Figures
Similar articles
-
Inhibition of NADH-ubiquinone reductase activity by N,N'-dicyclohexylcarbodiimide and correlation of this inhibition with the occurrence of energy-coupling site 1 in various organisms.Biochemistry. 1987 May 19;26(10):2822-8. doi: 10.1021/bi00384a025. Biochemistry. 1987. PMID: 3111526
-
Identification of the dicyclohexylcarbodiimide-binding subunit of NADH-ubiquinone oxidoreductase (Complex I).J Biol Chem. 1988 Nov 5;263(31):16150-5. J Biol Chem. 1988. PMID: 3141400
-
Mitochondrial nicotinamide nucleotide transhydrogenase: nonidentical modification by N,N'-dicyclohexylcarbodiimide and N-(ethoxycarbonyl)-2-ethoxy-1,2-dihydroquinoline at the NAD(H) binding site.Arch Biochem Biophys. 1985 Nov 15;243(1):298-304. doi: 10.1016/0003-9861(85)90799-4. Arch Biochem Biophys. 1985. PMID: 4062305
-
The Na+-translocating NADH:quinone oxidoreductase (NDH I) from Klebsiella pneumoniae and Escherichia coli: implications for the mechanism of redox-driven cation translocation by complex I.J Bioenerg Biomembr. 2001 Jun;33(3):179-86. doi: 10.1023/a:1010774701327. J Bioenerg Biomembr. 2001. PMID: 11695827 Review.
-
Na(+) translocation by bacterial NADH:quinone oxidoreductases: an extension to the complex-I family of primary redox pumps.Biochim Biophys Acta. 2001 May 1;1505(1):45-56. doi: 10.1016/s0005-2728(00)00276-0. Biochim Biophys Acta. 2001. PMID: 11248188 Review.
Cited by
-
Critical roles of subunit NuoH (ND1) in the assembly of peripheral subunits with the membrane domain of Escherichia coli NDH-1.J Biol Chem. 2009 Apr 10;284(15):9814-23. doi: 10.1074/jbc.M809468200. Epub 2009 Feb 3. J Biol Chem. 2009. PMID: 19189973 Free PMC article.
-
Essential regions in the membrane domain of bacterial complex I (NDH-1): the machinery for proton translocation.J Bioenerg Biomembr. 2014 Aug;46(4):279-87. doi: 10.1007/s10863-014-9558-8. Epub 2014 Jun 29. J Bioenerg Biomembr. 2014. PMID: 24973951 Review.
-
Cloning and Organelle Expression of Bamboo Mitochondrial Complex I Subunits Nad1, Nad2, Nad4, and Nad5 in the Yeast Saccharomyces cerevisiae.Int J Mol Sci. 2022 Apr 6;23(7):4054. doi: 10.3390/ijms23074054. Int J Mol Sci. 2022. PMID: 35409414 Free PMC article.
References
-
- Beinert, H. 1978. Micro methods for the quantitative determination of iron and copper in biological material. Methods Enzymol. 54:435-445. - PubMed
-
- Beinert, H. 1983. Semi-micro methods for analysis of labile sulfide and of labile sulfide plus sulfane sulfur in unusually stable iron-sulfur proteins. Anal. Biochem. 131:373-378. - PubMed
-
- Bertsova, Y. V., and A. V. Bogachev. 2004. The origin of the sodium-dependent NADH oxidation by the respiratory chain of Klebsiella pneumoniae. FEBS Lett. 563:207-212. - PubMed
-
- Böttcher, B., D. Scheide, M. Hesterberg, L. Nagel-Steger, and T. Friedrich. 2002. A novel, enzymatically active conformation of the Escherichia coli NADH:ubiquinone oxidoreductase (complex I). J. Biol. Chem. 277:17970-17977. - PubMed
-
- Carroll, J., I. M. Fearnley, J. M. Skehel, M. J. Runswick, R. J. Shannon, J. Hirst, and J. E. Walker. 2005. The post-translational modifications of the nuclear encoded subunits of complex I from bovine heart mitochondria. Mol. Cell. Proteomics 4:693-699. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
