

Mutational analysis of the chemoreceptor-coupling domain of the Escherichia coli chemotaxis signaling kinase CheA
- PMID: 16621823
- PMCID: PMC1447439
- DOI: 10.1128/JB.188.9.3299-3307.2006
Mutational analysis of the chemoreceptor-coupling domain of the Escherichia coli chemotaxis signaling kinase CheA
Abstract
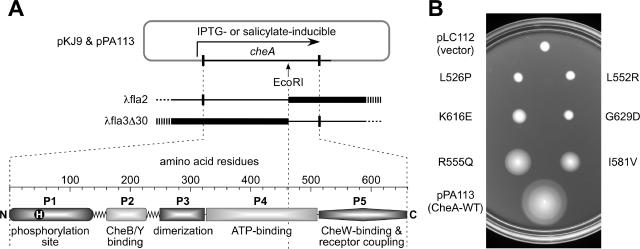
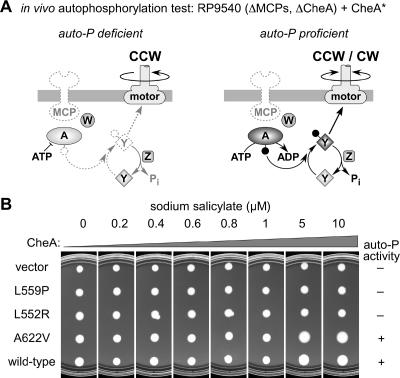
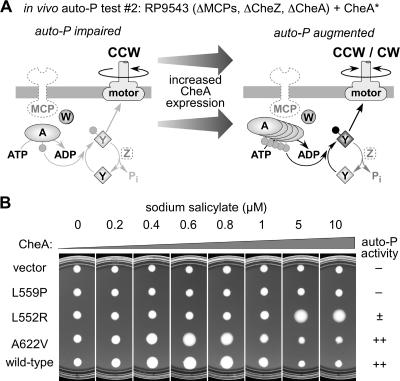
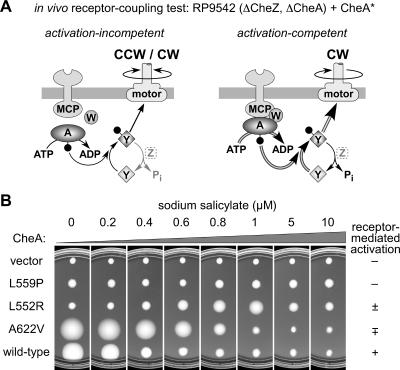
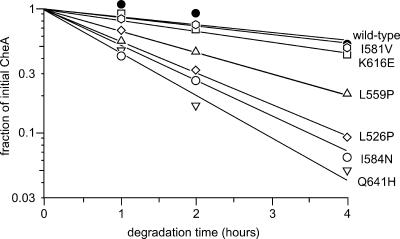
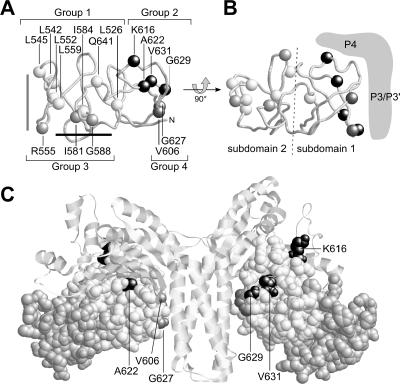
During chemotactic signaling by Escherichia coli, autophosphorylation of the histidine kinase CheA is coupled to chemoreceptor control by the CheW protein, which interacts with the C-terminal P5 domain of CheA. To identify P5 determinants important for CheW binding and receptor coupling control, we isolated and characterized a series of P5 missense mutants. The mutants fell into four phenotypic groups on the basis of in vivo behavioral and protein stability tests and in vitro assays with purified mutant proteins. Group 1 mutants exhibited autophosphorylation and receptor-coupling defects, and their CheA proteins were subject to relatively rapid degradation in vivo. Group 1 mutations were located at hydrophobic residues in P5 subdomain 2 and most likely caused folding defects. Group 2 mutants made stable CheA proteins with normal autophosphorylation ability but with defects in CheW binding and in receptor-mediated activation of CheA autophosphorylation. Their mutations affected residues in P5 subdomain 1 near the interface with the CheA dimerization (P3) and ATP-binding (P4) domains. Mutant proteins of group 3 were normal in all tests yet could not support chemotaxis, suggesting that P5 has one or more important but still unknown signaling functions. Group 4 mutant proteins were specifically defective in receptor-mediated deactivation control. The group 4 mutations were located in P5 subdomain 1 at the P3/P3' interface. We conclude that P5 subdomain 1 is important for CheW binding and for receptor coupling control and that these processes may require substantial motions of the P5 domain relative to the neighboring P3 and P4 domains of CheA.
Figures
Similar articles
-
Cysteine-scanning analysis of the chemoreceptor-coupling domain of the Escherichia coli chemotaxis signaling kinase CheA.J Bacteriol. 2006 Jun;188(12):4321-30. doi: 10.1128/JB.00274-06. J Bacteriol. 2006. PMID: 16740938 Free PMC article.
-
Identification of a Kinase-Active CheA Conformation in Escherichia coli Chemoreceptor Signaling Complexes.J Bacteriol. 2019 Nov 5;201(23):e00543-19. doi: 10.1128/JB.00543-19. Print 2019 Dec 1. J Bacteriol. 2019. PMID: 31501279 Free PMC article.
-
Structure of the ternary complex formed by a chemotaxis receptor signaling domain, the CheA histidine kinase, and the coupling protein CheW as determined by pulsed dipolar ESR spectroscopy.Biochemistry. 2010 May 11;49(18):3824-41. doi: 10.1021/bi100055m. Biochemistry. 2010. PMID: 20355710 Free PMC article.
-
Bacterial chemotaxis coupling protein: Structure, function and diversity.Microbiol Res. 2019 Feb;219:40-48. doi: 10.1016/j.micres.2018.11.001. Epub 2018 Nov 6. Microbiol Res. 2019. PMID: 30642465 Review.
-
Comparative genomic and protein sequence analyses of a complex system controlling bacterial chemotaxis.Methods Enzymol. 2007;422:1-31. doi: 10.1016/S0076-6879(06)22001-9. Methods Enzymol. 2007. PMID: 17628132 Free PMC article. Review.
Cited by
-
Regulatory Role of an Interdomain Linker in the Bacterial Chemotaxis Histidine Kinase CheA.J Bacteriol. 2018 Apr 24;200(10):e00052-18. doi: 10.1128/JB.00052-18. Print 2018 May 15. J Bacteriol. 2018. PMID: 29483161 Free PMC article.
-
Signalling-dependent interactions between the kinase-coupling protein CheW and chemoreceptors in living cells.Mol Microbiol. 2014 Sep;93(6):1144-55. doi: 10.1111/mmi.12727. Epub 2014 Aug 5. Mol Microbiol. 2014. PMID: 25060668 Free PMC article.
-
Chemotaxis arrays in Vibrio species and their intracellular positioning by the ParC/ParP system.J Bacteriol. 2018 Mar 12;200(15):e00793-17. doi: 10.1128/JB.00793-17. Online ahead of print. J Bacteriol. 2018. PMID: 29531180 Free PMC article.
-
The 3.2 Å resolution structure of a receptor: CheA:CheW signaling complex defines overlapping binding sites and key residue interactions within bacterial chemosensory arrays.Biochemistry. 2013 Jun 4;52(22):3852-65. doi: 10.1021/bi400383e. Epub 2013 May 23. Biochemistry. 2013. PMID: 23668907 Free PMC article.
-
CheA-receptor interaction sites in bacterial chemotaxis.J Mol Biol. 2012 Sep 14;422(2):282-90. doi: 10.1016/j.jmb.2012.05.023. Epub 2012 May 30. J Mol Biol. 2012. PMID: 22659323 Free PMC article.
References
-
- Ames, P., Y. A. Yu, and J. S. Parkinson. 1996. Methylation segments are not required for chemotactic signalling by cytoplasmic fragments of Tsr, the methyl-accepting serine chemoreceptor of Escherichia coli. Mol. Microbiol. 19:737-746. - PubMed
-
- Bilwes, A. M., L. A. Alex, B. R. Crane, and M. I. Simon. 1999. Structure of CheA, a signal-transducing histidine kinase. Cell 96:131-141. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
