

Rtn1p is involved in structuring the cortical endoplasmic reticulum
- PMID: 16624861
- PMCID: PMC1483037
- DOI: 10.1091/mbc.e06-01-0080
Rtn1p is involved in structuring the cortical endoplasmic reticulum
Abstract
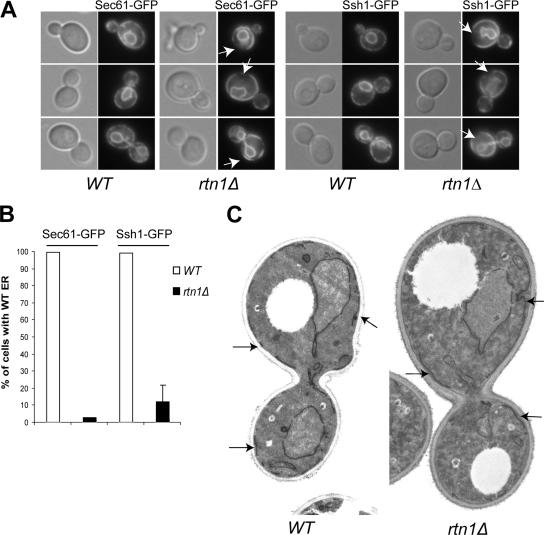
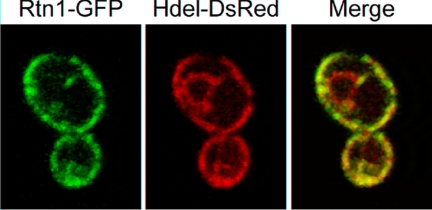
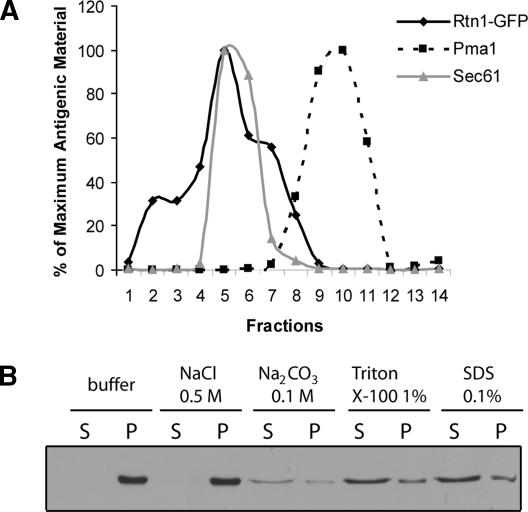
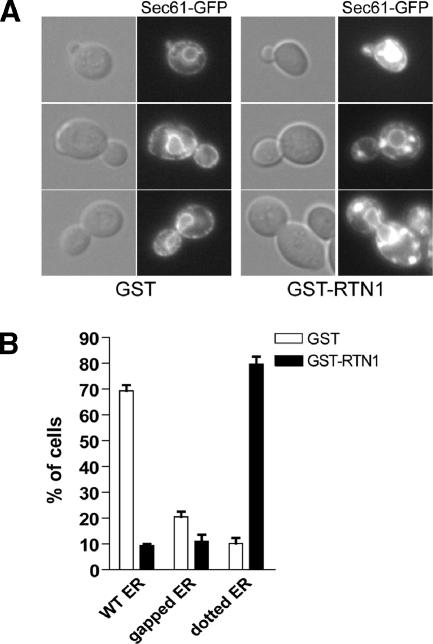
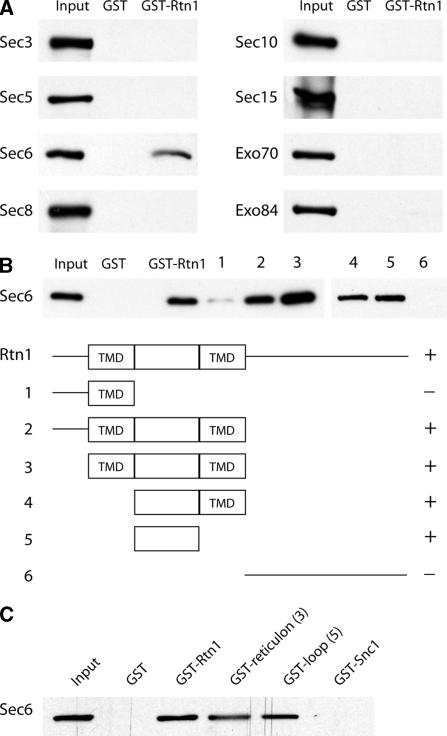
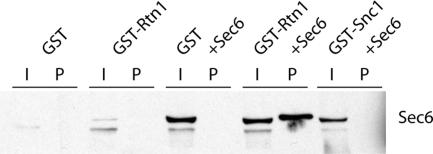
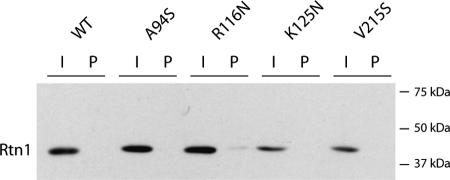
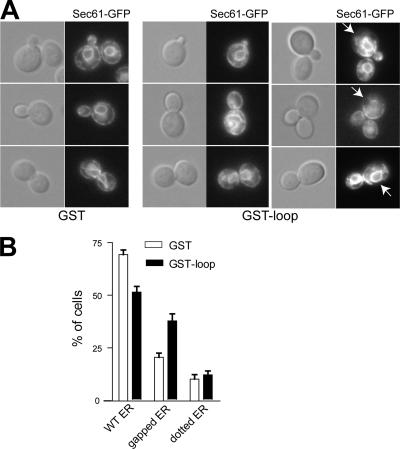
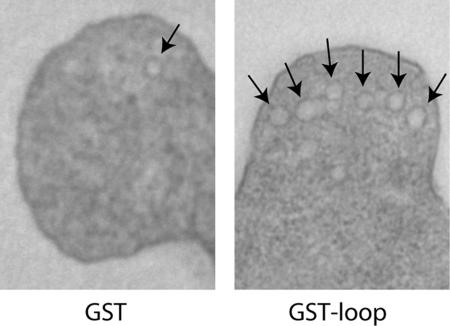
The endoplasmic reticulum (ER) contains both cisternal and reticular elements in one contiguous structure. We identified rtn1Delta in a systematic screen for yeast mutants with altered ER morphology. The ER in rtn1Delta cells is predominantly cisternal rather than reticular, yet the net surface area of ER is not significantly changed. Rtn1-green fluorescent protein (GFP) associates with the reticular ER at the cell cortex and with the tubules that connect the cortical ER to the nuclear envelope, but not with the nuclear envelope itself. Rtn1p overexpression also results in an altered ER structure. Rtn proteins are found on the ER in a wide range of eukaryotes and are defined by two membrane-spanning domains flanking a conserved hydrophilic loop. Our results suggest that Rtn proteins may direct the formation of reticulated ER. We independently identified Rtn1p in a proteomic screen for proteins associated with the exocyst vesicle tethering complex. The conserved hydophilic loop of Rtn1p binds to the exocyst subunit Sec6p. Overexpression of this loop results in a modest accumulation of secretory vesicles, suggesting impaired exocyst function. The interaction of Rtn1p with the exocyst at the bud tip may trigger the formation of a cortical ER network in yeast buds.
Figures

References
-
- Baumann O., Walz B. Endoplasmic reticulum of animal cells and its organization into structural and functional domains. Int. Rev. Cytol. 2001;205:149–214. - PubMed
-
- Chen M. S., Huber A. B., van der Haar M. E., Frank M., Schnell L., Spillmann A. A., Christ F., Schwab M. E. Nogo-A is a myelin-associated neurite outgrowth inhibitor and an antigen for mAb IN-1. Nature. 2000;403:434–439. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
