

Light-induced changes in spike synchronization between coupled ON direction selective ganglion cells in the mammalian retina
- PMID: 16624941
- PMCID: PMC6673999
- DOI: 10.1523/JNEUROSCI.0496-06.2006
Light-induced changes in spike synchronization between coupled ON direction selective ganglion cells in the mammalian retina
Abstract
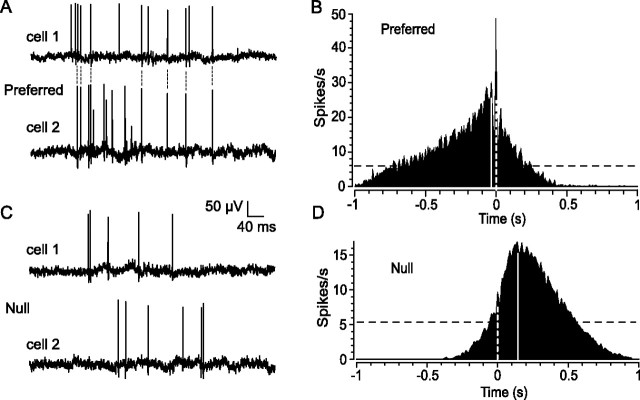
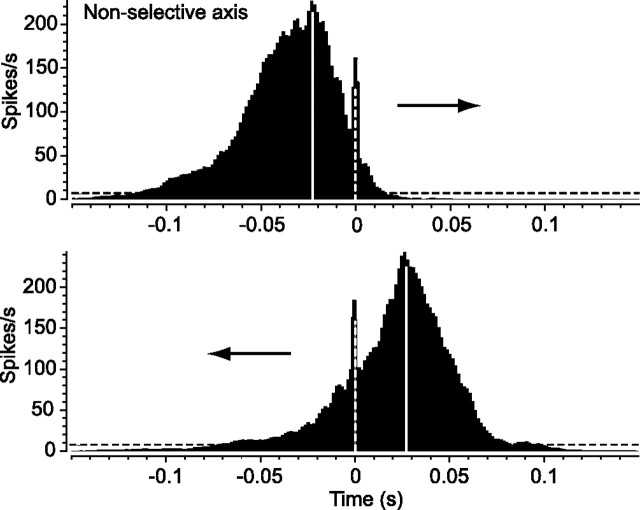
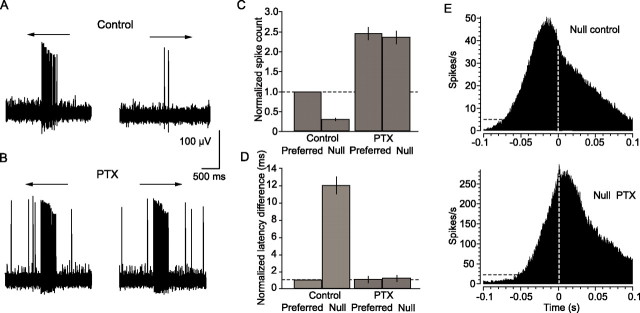
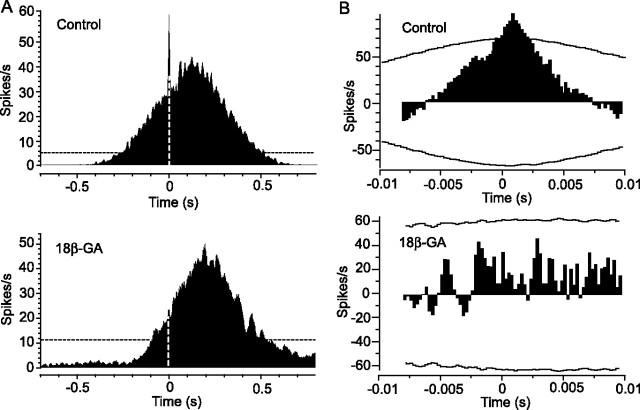
Although electrical coupling via gap junctions is prevalent among ganglion cells in the vertebrate retina, there have been few direct studies of their influence on the light-evoked signaling of these cells. Here, we describe the pattern and function of coupling between the ON direction selective (DS) ganglion cells, a unique subtype whose signals are transmitted to the accessory optic system (AOS) where they initiate the optokinetic response. ON DS cells are coupled indirectly via gap junctions made with a subtype of polyaxonal amacrine cell. This coupling underlies synchronization of the spontaneous and light-evoked spike activity of neighboring ON DS cells. However, we find that ON DS cell pairs show robust synchrony for all directions of stimulus movement, except for the null direction. Null stimulus movement evokes a GABAergic inhibition that temporally shifts firing of ON DS cell neighbors, resulting in a desynchronization of spike activity. Thus, detection of null stimulus movement appears key to the direction selectivity of ON DS cells, evoking both an attenuation of spike frequency and a desynchronization of neighbors. We posit that active desynchronization reduces summation of synaptic potentials at target AOS cells and thus provides a secondary mechanism by which ON DS cell ensembles can signal direction of stimulus motion to the brain.
Figures
References
-
- Alonso JM, Usrey WM, Reid RC (1996). Precisely correlated firing in cells of the lateral geniculate nucleus. Nature 383:815–819. - PubMed
-
- Amthor FR, Takahashi ES, Oyster CW (1989). Morphologies of rabbit retinal ganglion cells with complex receptive fields. J Comp Neurol 280:97–121. - PubMed
-
- Beyer EC, Paul DL, Goodenough DA (1990). Connexin family of gap junction proteins. J Membr Biol 116:87–194. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous