

Compartment-dependent colocalization of Kir3.2-containing K+ channels and GABAB receptors in hippocampal pyramidal cells
- PMID: 16624949
- PMCID: PMC6673994
- DOI: 10.1523/JNEUROSCI.4178-05.2006
Compartment-dependent colocalization of Kir3.2-containing K+ channels and GABAB receptors in hippocampal pyramidal cells
Abstract
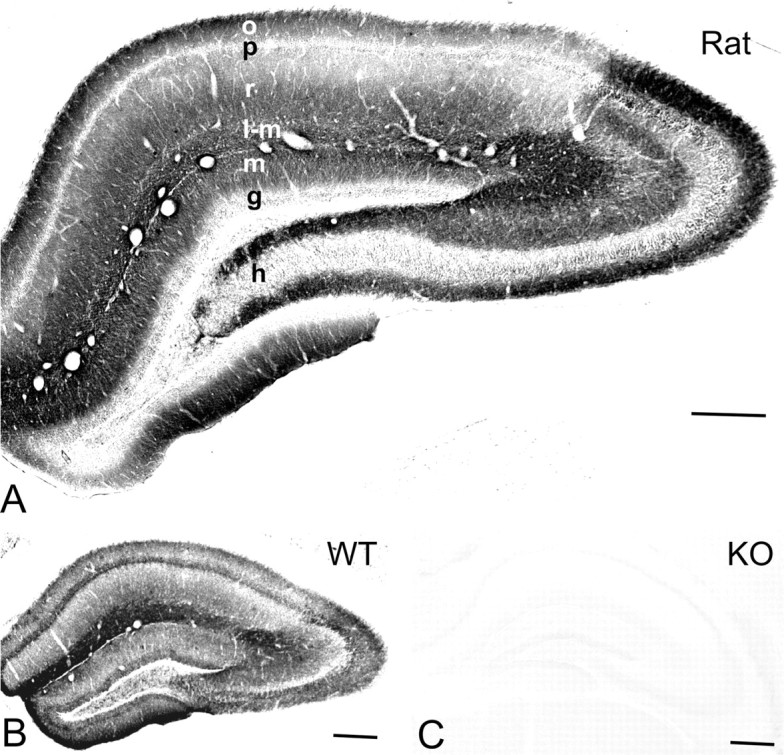
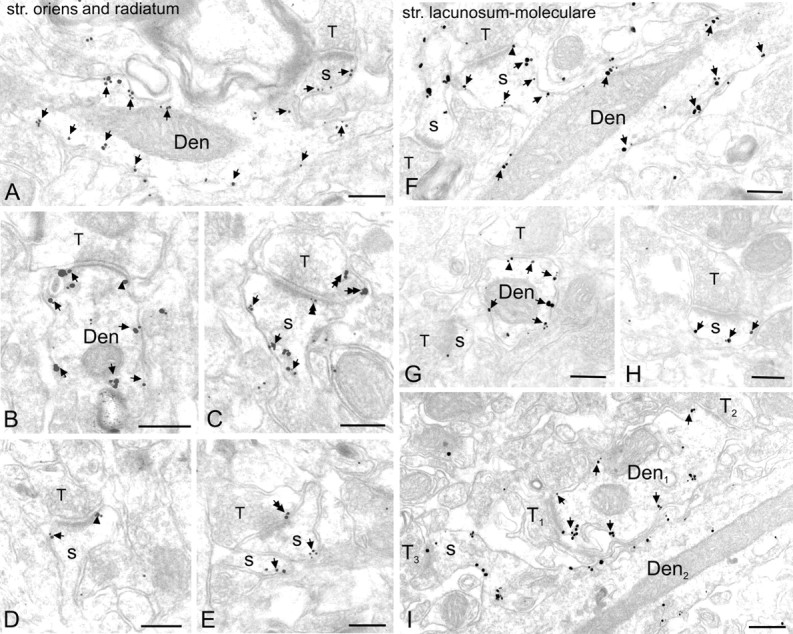
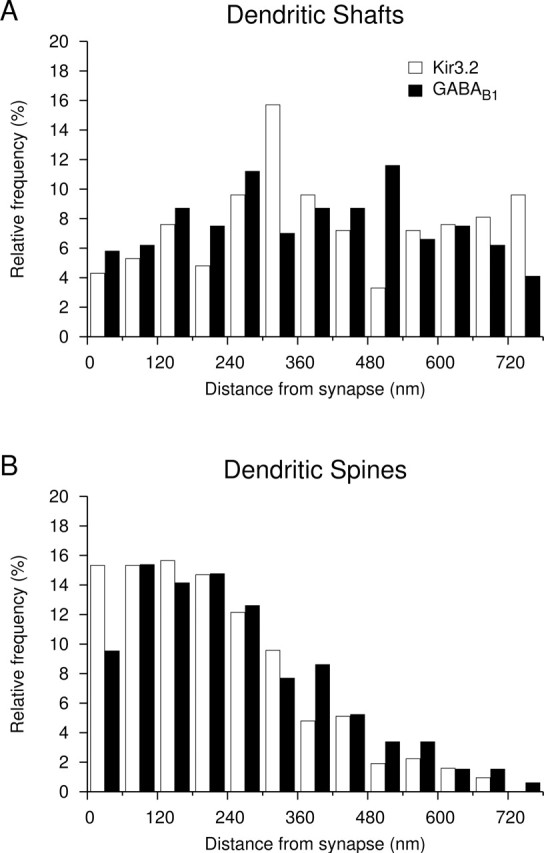
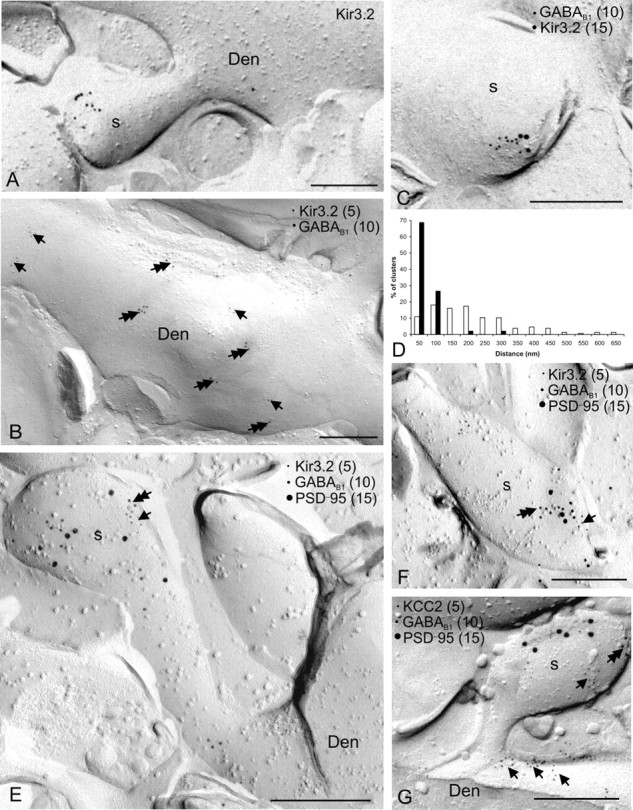
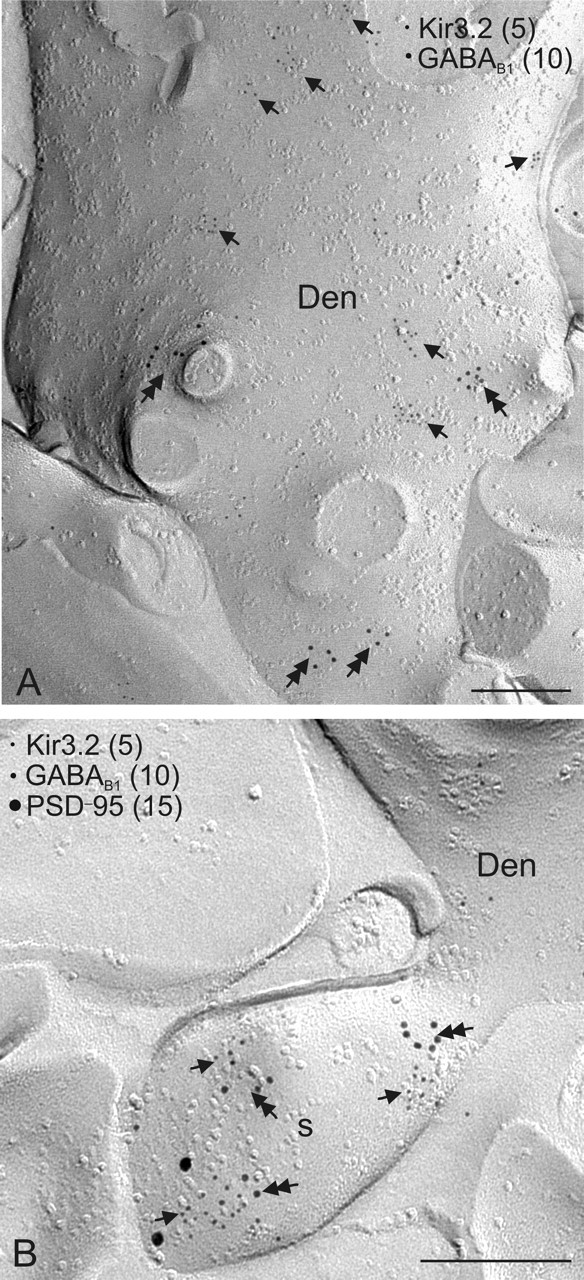
G-protein-coupled inwardly rectifying K+ channels (Kir3 channels) coupled to metabotropic GABAB receptors are essential for the control of neuronal excitation. To determine the distribution of Kir3 channels and their spatial relationship to GABAB receptors on hippocampal pyramidal cells, we used a high-resolution immunocytochemical approach. Immunoreactivity for the Kir3.2 subunit was most abundant postsynaptically and localized to the extrasynaptic plasma membrane of dendritic shafts and spines of principal cells. Quantitative analysis of immunogold particles for Kir3.2 revealed an enrichment of the protein around putative glutamatergic synapses on dendritic spines, similar to that of GABA(B1). Consistent with this observation, a high degree of coclustering of Kir3.2 and GABA(B1) was revealed around excitatory synapses by the highly sensitive SDS-digested freeze-fracture replica immunolabeling. In contrast, in dendritic shafts receptors and channels were found to be mainly segregated. These results suggest that Kir3.2-containing K+ channels on dendritic spines preferentially mediate the effect of GABA, whereas channels on dendritic shafts are likely to be activated by other neurotransmitters as well. Thus, Kir3 channels, localized to different subcellular compartments of hippocampal principal cells, appear to be differentially involved in synaptic integration in pyramidal cell dendrites.
Figures
References
-
- Andrade R, Malenka RC, Nicoll RA (1986). A G protein couples serotonin and GABAB receptors to the same channels in hippocampus. Science 234:1261–1265. - PubMed
-
- Bichet D, Haass FA, Jan LY (2003). Merging functional studies with structures of inward rectifier K+ channels. Nat Rev Neurosci 4:957–967. - PubMed
-
- Cryan JF, Kaupmann K (2005). Don’t worry “B” happy!: a role for GABAB receptors in anxiety and depression. Trends Pharmacol Sci 26:36–43. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources