

Review
doi: 10.1038/nrm1925.
Lipid rafts: contentious only from simplistic standpoints
Affiliations
- PMID: 16625153
- PMCID: PMC2782566
- DOI: 10.1038/nrm1925
Item in Clipboard
Review
Lipid rafts: contentious only from simplistic standpoints
Nat Rev Mol Cell Biol.
2006 Jun.
Abstract
The hypothesis that lipid rafts exist in plasma membranes and have crucial biological functions remains controversial. The lateral heterogeneity of proteins in the plasma membrane is undisputed, but the contribution of cholesterol-dependent lipid assemblies to this complex, non-random organization promotes vigorous debate. In the light of recent studies with model membranes, computational modelling and innovative cell biology, I propose an updated model of lipid rafts that readily accommodates diverse views on plasma-membrane micro-organization.
Figures
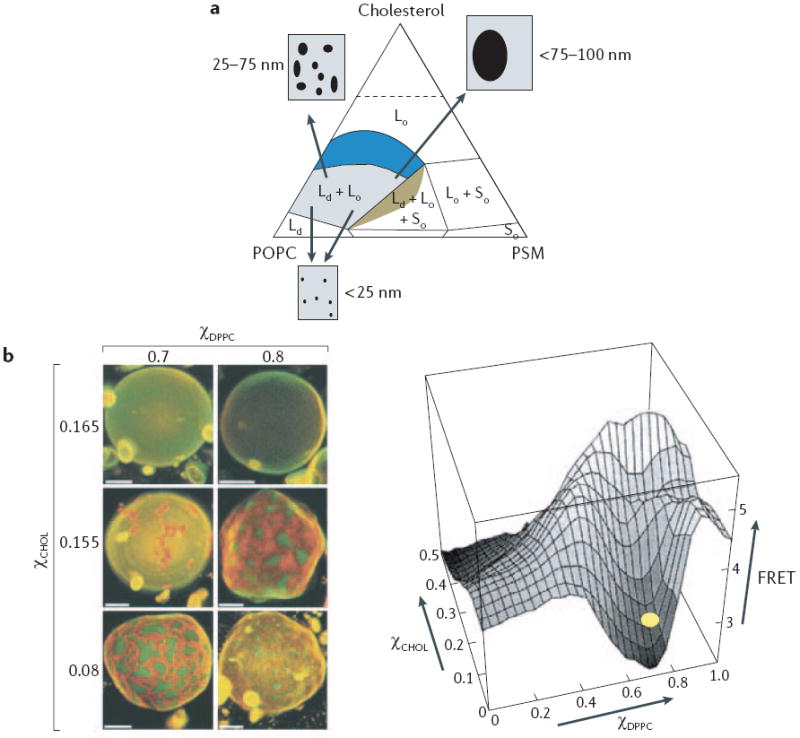
a Phase diagram at 23°C for ternary mixtures of cholesterol, sphingomyelin (PSM) and l-palmitoyl-2-oleoyl-phosphatidylcholine (POPC). Each vertex of the diagram corresponds to a 100% content of each lipid. Coloured regions represent membrane compositions that can form a liquid disordered (Ld) phase. Within the region of liquid ordered (Lo)–Ld co-existence (blue), varying the cholesterol percentages from ~10% to 35% progressively increases the size of Lo domains that are detected by fluorescence resonance energy transfer (FRET) imaging or standard microscopy. It is important to note that phase diagrams such as this are crucially dependent on temperature, although an extensive area of Lo–Ld phase co-existence is present at 37°C(see REFS 3,6). b Macroscopic domain formation in giant unilamellar vesicles that are composed of cholesterol, 1,2-dipalmitoylphosphatidylcholine (DPPC) and 1,2-dioleoylphosphatidylcholine (DOPC) and visualized using dyes that preferentially partition into Lo domains (orange) and Ld domains (green). When the cholesterol percentage (χCHOL) is >16%, macroscopic domain formation is no longer seen, but nanodomain formation is still detectable as a loss of a FRET signal between two probes that preferentially partition into Lo and Ld domains. The yellow oval marks the approximate composition of the top row of vesicles. Scale bar = 5μm. χDPPC = DPPC content as a fraction of DPPC and DOPC. Part a is reproduced with permission from REF. © (2005) Elsevier; part b is reproduced with permission from REF. © (2001) The Biophysical Society.
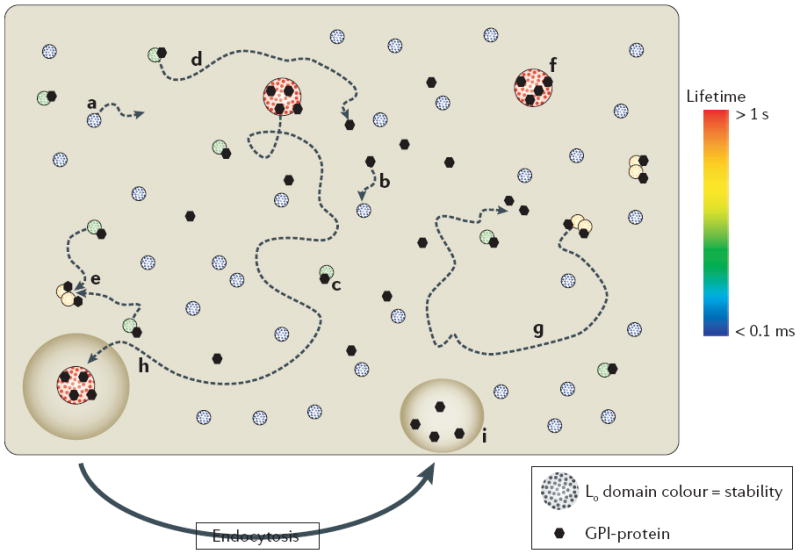
Liquid ordered (Lo) domains in the plasma membrane are heterogeneous in size and lifetime (from >1 s to <0.1 ms, as indicated by colour). Lo domain stability or lifetime is a function of size, capture by a raft-stabilizing protein and protein–protein interactions of constituent proteins. The length of trajectories (dotted arrowed lines) and, therefore, the probability of collision with proteins or other Lo domains are proportional to lifetime. For simplicity, only sample trajectories are shown — however, all of the domains and proteins that are shown should be envisaged to diffuse laterally. The diagram illustrates the fate of different classes of Lo domain. Small, unstable Lo domains form spontaneously, diffuse laterally in the plasma membrane, but have a limited lifetime (a). If captured by a glycosylphosphatidylinisotol (GPI)-anchored protein (or other raft-stabilizing proteins) (b) the stability of the Lo domain is increased with the formation of a complex (c), which has two possible outcomes: the complex diffuses laterally but the Lo domain disassembles (d), or there is a collision with other protein-stabilized Lo domains that creates protein clusters (e). Further collisions will generate larger, more stable Lo–protein complexes (f) that are further stabilized by protein–protein interactions, or in the absence of collisions (g) the Lo–protein complex disassembles. The fate of larger, more stable Lo–protein complexes (h) is capture by endocytic pathways that disassemble the complexes and return lipid and protein constituents back to the plasma membrane (i). Intuitively, the model predicts the generation of protein clusters that is dependent on their interaction with Lo domains. Endocytosis indicates endocytosis that is specific for larger raft complexes (as shown), possibly all endocytic processes contribute to limiting the size of raft domains. Note that the larger, more stable lipid–protein complexes (f) could operate the way rafts are proposed to in the classic model, with dynamic partitioning of proteins between the domain and the surrounding disordered membrane (not shown).
References
-
- Simons K, Ikonen E. Functional rafts in cell membranes. Nature. 1997;387:569–572. - PubMed
-
- Edidin M. The state of lipid rafts: from model membranes to cells. Annu Rev Biophys Biomol Struct. 2003;32:257–283. - PubMed
-
- Simons K, Vaz WL. Model systems, lipid rafts, and cell membranes. Anna Rev Biophys Biomol Struct. 2004;33:269–295. - PubMed
-
- Munro S. Lipid rafts: elusive or illusive? Cell. 2003;115:377–388. - PubMed
-
- Nichols B. Cell biology: without a raft. Nature. 2005;436:638–639. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
