

Tmc1 is necessary for normal functional maturation and survival of inner and outer hair cells in the mouse cochlea
- PMID: 16627570
- PMCID: PMC1817746
- DOI: 10.1113/jphysiol.2005.095661
Tmc1 is necessary for normal functional maturation and survival of inner and outer hair cells in the mouse cochlea
Abstract
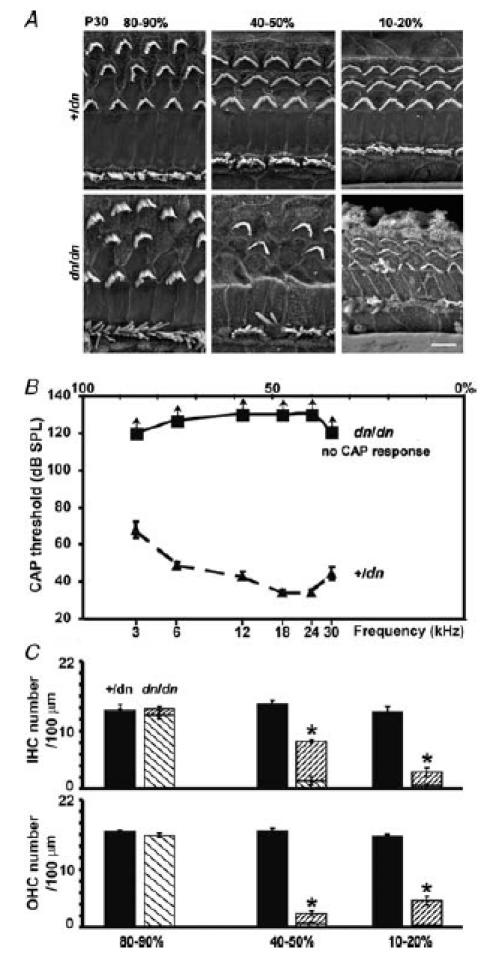
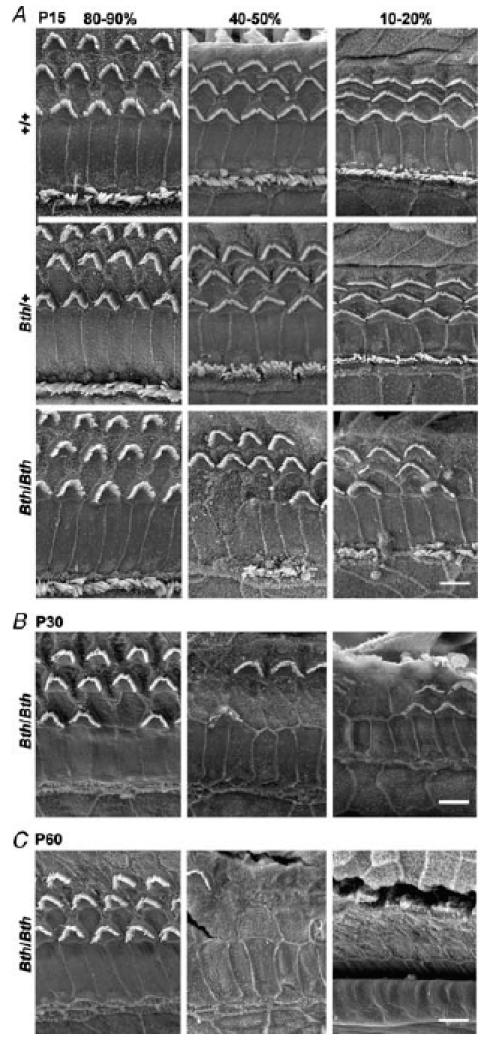
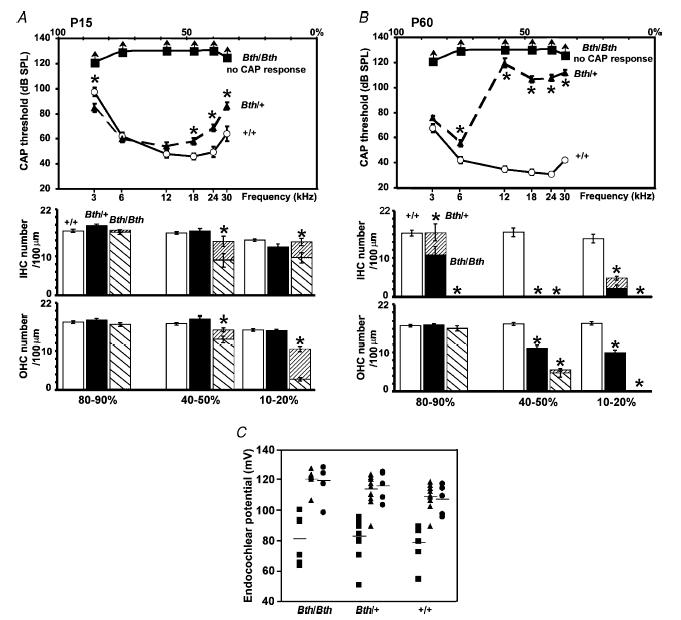
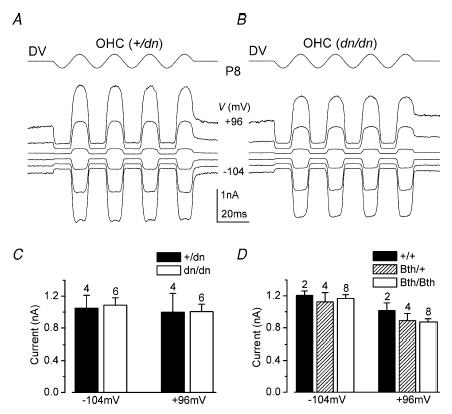
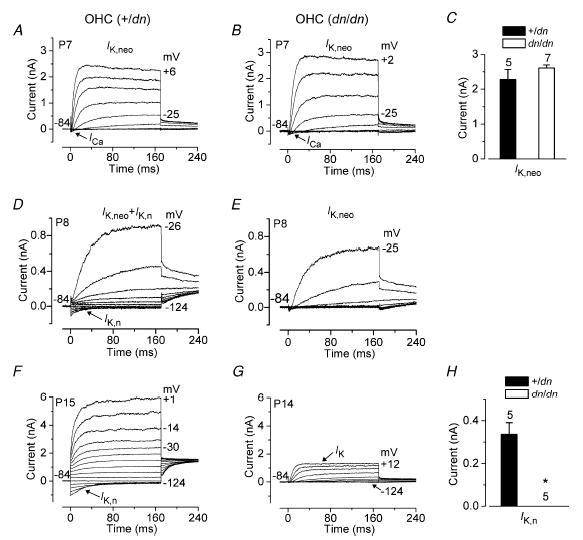
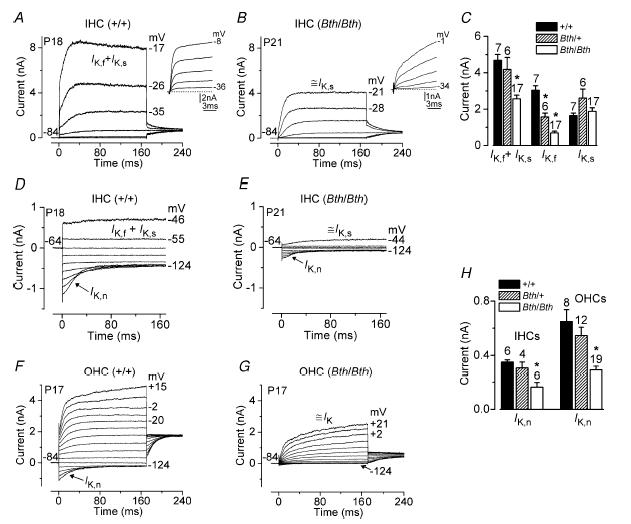
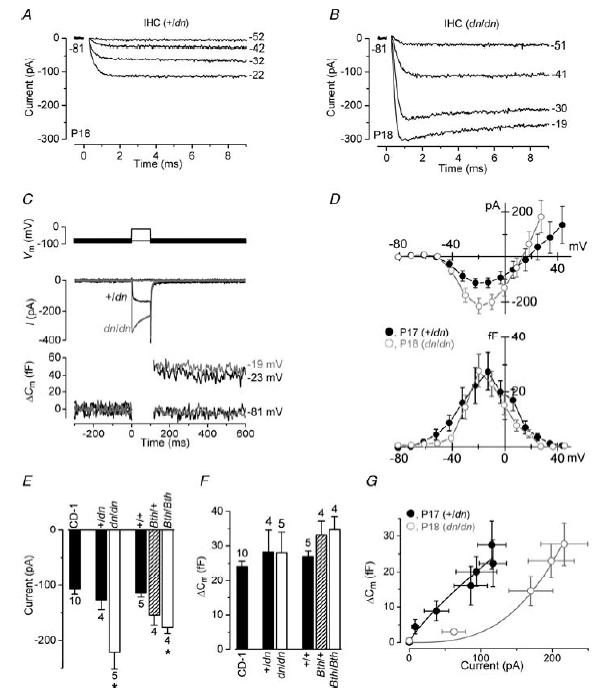
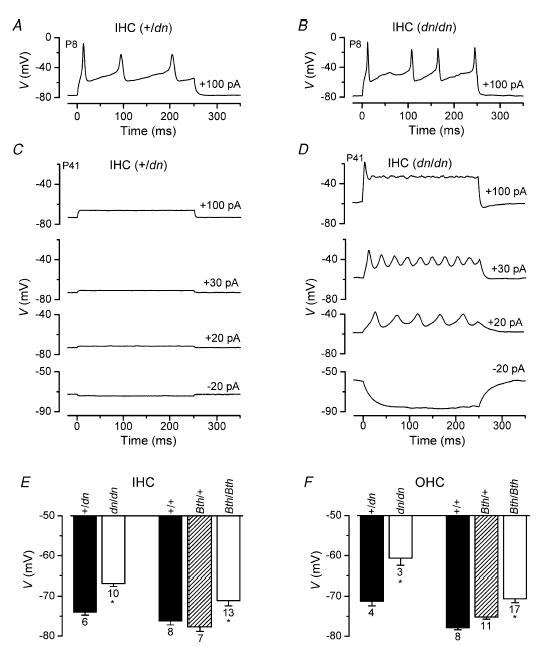
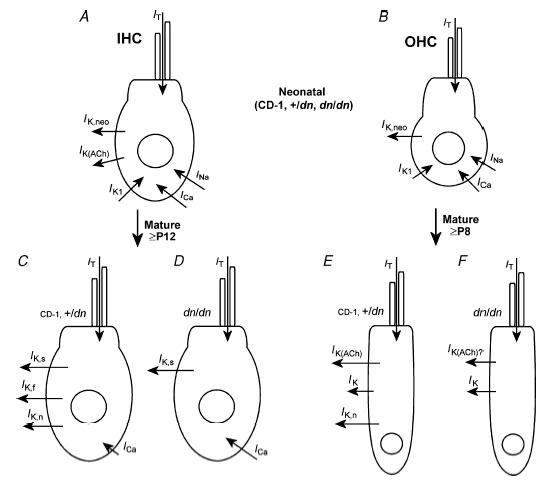
The deafness (dn) and Beethoven (Bth) mutant mice are models for profound congenital deafness (DFNB7/B11) and progressive hearing loss (DFNA36), respectively, caused by recessive and dominant mutations of transmembrane cochlear-expressed gene 1 (TMC1), which encodes a transmembrane protein of unknown function. In the mouse cochlea Tmc1 is expressed in both outer (OHCs) and inner (IHCs) hair cells from early stages of development. Immature hair cells of mutant mice seem normal in appearance and biophysical properties. From around P8 for OHCs and P12 for IHCs, mutants fail to acquire (dn/dn) or show reduced expression (Bth/Bth and, to a lesser extent Bth/+) of the K+ currents which contribute to their normal functional maturation (the BK-type current IK,f in IHCs, and the delayed rectifier IK,n in both cell types). Moreover, the exocytotic machinery in mutant IHCs does not develop normally as judged by the persistence of immature features of the Ca2+ current and exocytosis into adulthood. Mutant mice exhibited progressive hair cell damage and loss. The compound action potential (CAP) thresholds of Bth/+ mice were raised and correlated with the degree of hair cell loss. Homozygous mutants (dn/dn and Bth/Bth) never showed CAP responses, even at ages where many hair cells were still present in the apex of the cochlea, suggesting their hair cells never function normally. We propose that Tmc1 is involved in trafficking of molecules to the plasma membrane or serves as an intracellular regulatory signal for differentiation of immature hair cells into fully functional auditory receptors.
Figures

References
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous