

Crossbridge and non-crossbridge contributions to tension in lengthening rat muscle: force-induced reversal of the power stroke
- PMID: 16627571
- PMCID: PMC1779750
- DOI: 10.1113/jphysiol.2005.095448
Crossbridge and non-crossbridge contributions to tension in lengthening rat muscle: force-induced reversal of the power stroke
Abstract
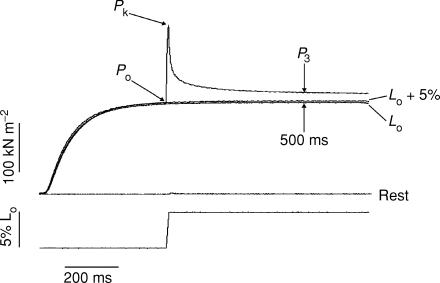
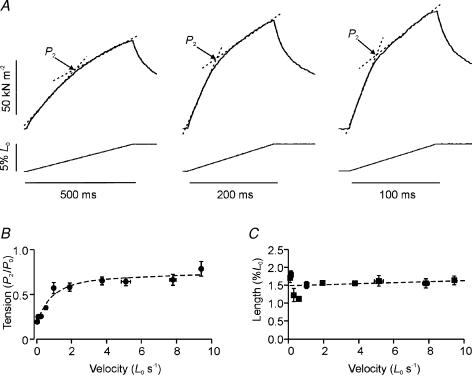
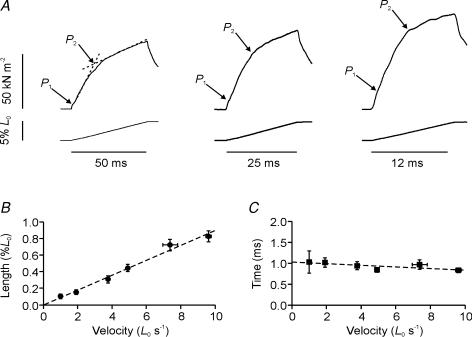
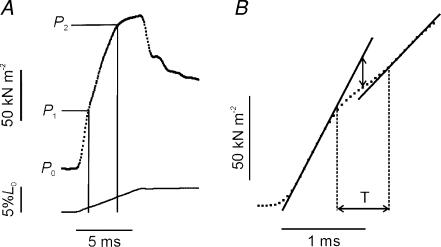
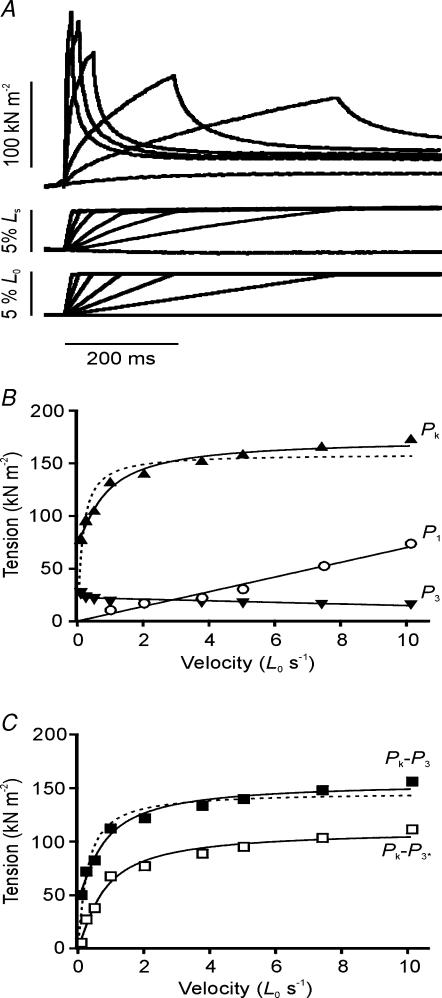
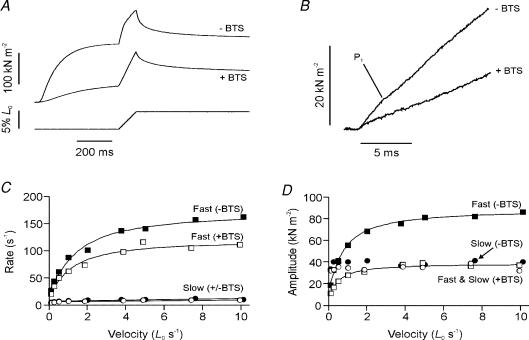
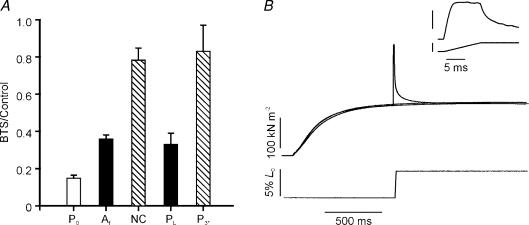
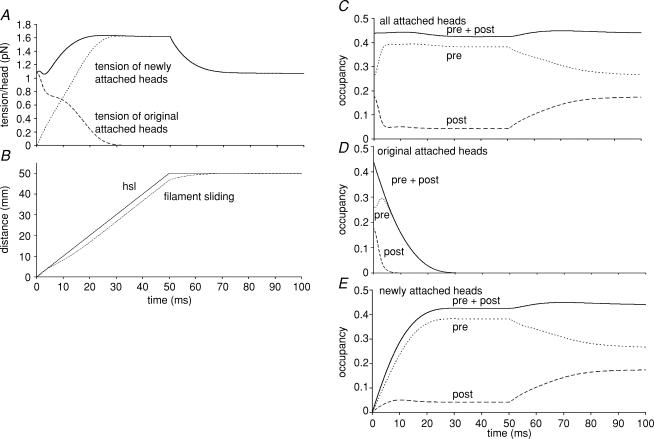
Lengthening of active muscle is an essential feature of animal locomotion, but the molecular processes occurring are incompletely understood. We therefore examined and modelled tension responses to ramp stretches (5% fibre length, L0) over a wide range of velocities (0.1-10 L(0) s(-1)) of tetanized intact rat muscle fibre bundles (L0 approximately 2 mm) with a resting sarcomere length of 2.5 microm at 20 degrees C. Tension rose to a peak during stretch and decayed afterwards to a level which was higher than the prestretch tetanic tension. This residual force enhancement was insensitive to velocity. The tension rise during stretch showed an early transition (often appearing as an inflection) at approximately 1 ms. Both the stretch (L1) and the tension rise at this transition increased in proportion to velocity. A second transition, marked by a reduction in slope, occurred at a stretch of approximately 18 nm per half-sarcomere; the rise in tension at this transition increased with velocity towards a plateau. Based on analyses of the velocity dependence of the tension and modelling, we propose that the initial steep increase in tension arises from increasing strain of all attached crossbridges and that the first transition reflects the tension loss due to the original post-stroke heads executing a reverse power stroke. Modelling indicates that the reduction in slope at the second transition occurs when the last of the heads that were attached at the start of the ramp become detached. Thereafter, the crossbridge cycle is largely truncated, with prepower stroke crossbridges rapidly detaching at high strain and attaching at low strain, the tension being borne mainly by the prestroke heads. Analysis of the tension decay after the ramp and the velocity dependence of the peak tension suggest that a non-crossbridge component increasingly develops tension throughout the stretch; this decays only slowly, reaching at 500 ms after the ramp approximately 20% of its peak value. This is supported by the finding that, in the presence of 10 microm N-benzyl-p-toluene sulphonamide (a myosin inhibitor), while isometric tension is reduced to approximately 15%, and the crossbridge contribution to stretch-induced tension rise is reduced to 30-40%, the peak non-crossbridge contribution and the residual force enhancement remain high. We propose that the residual force enhancement is due to changes upon activation in parallel elastic elements, specifically that titin stiffens and C-protein-actin interactions may be recruited.
Figures
Similar articles
-
Comparison of the tension responses to ramp shortening and lengthening in intact mammalian muscle fibres: crossbridge and non-crossbridge contributions.J Muscle Res Cell Motil. 2007;28(2-3):123-39. doi: 10.1007/s10974-007-9110-0. Epub 2007 Jul 4. J Muscle Res Cell Motil. 2007. PMID: 17610136
-
Crossbridge and non-crossbridge contributions to force in shortening and lengthening muscle.Adv Exp Med Biol. 2010;682:207-21. doi: 10.1007/978-1-4419-6366-6_12. Adv Exp Med Biol. 2010. PMID: 20824528 Review.
-
The visco-elasticity of resting intact mammalian (rat) fast muscle fibres.J Muscle Res Cell Motil. 1996 Jun;17(3):357-64. doi: 10.1007/BF00240933. J Muscle Res Cell Motil. 1996. PMID: 8814555
-
Crossbridge properties investigated by fast ramp stretching of activated frog muscle fibres.J Physiol. 2005 May 15;565(Pt 1):261-8. doi: 10.1113/jphysiol.2005.085209. Epub 2005 Mar 17. J Physiol. 2005. PMID: 15774512 Free PMC article.
-
Crossbridge and filament compliance in muscle: implications for tension generation and lever arm swing.J Muscle Res Cell Motil. 2010 Dec;31(4):245-65. doi: 10.1007/s10974-010-9232-7. Epub 2010 Dec 4. J Muscle Res Cell Motil. 2010. PMID: 21132353 Review.
Cited by
-
Pre-power-stroke cross-bridges contribute to force transients during imposed shortening in isolated muscle fibers.PLoS One. 2012;7(1):e29356. doi: 10.1371/journal.pone.0029356. Epub 2012 Jan 5. PLoS One. 2012. PMID: 22242168 Free PMC article.
-
Force-generating cross-bridges during ramp-shaped releases: evidence for a new structural state.Biophys J. 2009 Feb 18;96(4):1430-46. doi: 10.1016/j.bpj.2008.11.023. Biophys J. 2009. PMID: 19217860 Free PMC article.
-
Force enhancement in lengthening contractions of cat soleus muscle in situ: transient and steady-state aspects.Physiol Rep. 2013 Jul;1(2):e00017. doi: 10.1002/phy2.17. Epub 2013 Jun 28. Physiol Rep. 2013. PMID: 24303106 Free PMC article.
-
Importance of contraction history on muscle force of porcine urinary bladder smooth muscle.Int Urol Nephrol. 2017 Feb;49(2):205-214. doi: 10.1007/s11255-016-1482-x. Epub 2016 Dec 17. Int Urol Nephrol. 2017. PMID: 27988910
-
Millisecond-scale biochemical response to change in strain.Biophys J. 2011 Nov 16;101(10):2445-54. doi: 10.1016/j.bpj.2011.10.007. Epub 2011 Nov 15. Biophys J. 2011. PMID: 22098743 Free PMC article.
References
-
- Abbott BC, Aubert XM. Changes of energy in a muscle during very slow stretches. Proc R Soc Lond B. 1951;139:104–117. - PubMed
-
- Bagni MA, Cecchi G, Cecchini E, Colombini B, Colomo F. Force responses to fast ramp stretches in stimulated frog skeletal muscle fibres. J Muscle Res Cell Motil. 1998;19:33–42. - PubMed
-
- Brenner B. Mechanical and structural approaches to correlation of cross-bridge action in muscle with actomyosin ATPase in solution. Ann Rev Physiol. 1987;49:655–672. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources