

Breast milk lactoferrin regulates gene expression by binding bacterial DNA CpG motifs but not genomic DNA promoters in model intestinal cells
- PMID: 16627877
- PMCID: PMC3223379
- DOI: 10.1203/01.pdr.0000214958.80011.e1
Breast milk lactoferrin regulates gene expression by binding bacterial DNA CpG motifs but not genomic DNA promoters in model intestinal cells
Abstract
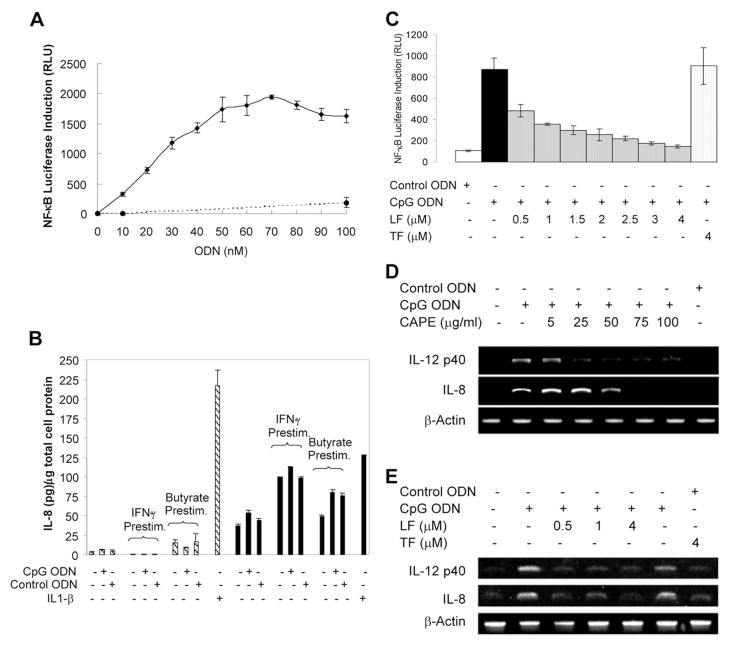
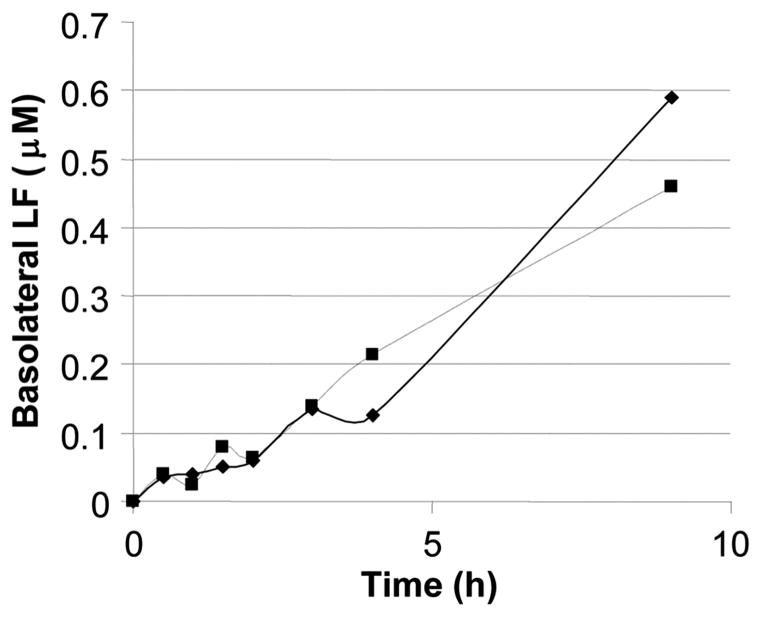
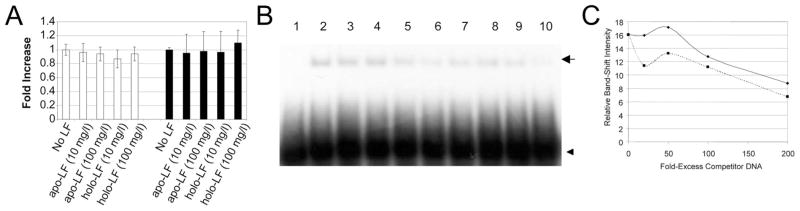
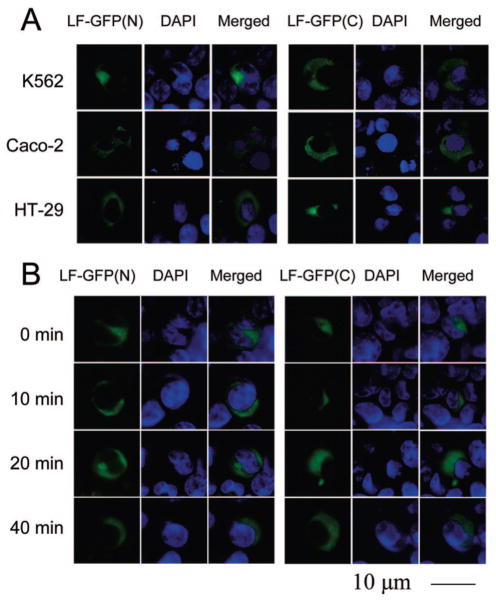
High-affinity binding of DNA by lactoferrin (LF) is an established phenomenon, but the biologic function of this interaction remains unclear. LF is an abundant breast milk protein (12.5-87.5 micromol/L) and is resistant to digestion in the infant gut. Regulation of gene expression by LF appears to be a major activity, particularly in modulating immune responses. We hypothesized that LF binding to DNA is a mechanism of gene regulation and aimed to identify the mechanism and physiologic sites of this activity. Our studies focused on two major biologic compartments of DNA: LF binding to proinflammatory bacterial DNA sequences (CpG motifs) in extracellular compartments and LF binding to genomic DNA promoters in the nucleus. LF 0.5 mmol/L inhibited CpG motif-induced nuclear factor-kappaB (NF-kappaB) activation and interleukin (IL)-8 and IL-12 cytokine gene transcription in B cells. Intestinal epithelial cells were unresponsive to CpG motifs. However, significant LF transferred across M cell-like monolayers, specialized epithelial cells that transcytose intact macromolecules to underlying B-cell follicles in the intestine. LF did not activate gene expression by binding to putative response elements in epithelial and lymphoid cells. Nor did LF bind to putative response elements specifically in gel-shift assays. No nuclear localization of LF was detected in green fluorescent protein (GFP) tagging experiments. We conclude that breast milk LF regulates gene expression by binding CpG motifs extracellularly, with follicular B cells in the infant intestine a likely target.
Figures
References
-
- Querinjean P, Masson PL, Heremans JF. Molecular weight, single-chain structure and amino acid composition of human lactoferrin. Eur J Biochem. 1971;20:420–425. - PubMed
-
- Brock J. Lactoferrin: a multifunctional immunoregulatory protein? Immunol Today. 1995;16:417–419. - PubMed
-
- Lonnerdal B, Adkins Y. Developmental changes in breast milk protein composition during lactation. In: Sanderson IR, Walker WA, editors. Development of the Gastrointestinal Tract. BC Decker Inc; Hamilton: 2000. pp. 227–244.
-
- Brines RD, Brock JH. The effect of trypsin and chymotrypsin on the in vitro antimicrobial and iron-binding properties of lactoferrin in human milk and bovine colostrum. Unusual resistance of human apolactoferrin to proteolytic digestion. Biochim Biophys Acta. 1983;759:229–235. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Miscellaneous
