

A conserved pathway to activate BRCA1-dependent ubiquitylation at DNA damage sites
- PMID: 16628214
- PMCID: PMC1462971
- DOI: 10.1038/sj.emboj.7601102
A conserved pathway to activate BRCA1-dependent ubiquitylation at DNA damage sites
Abstract
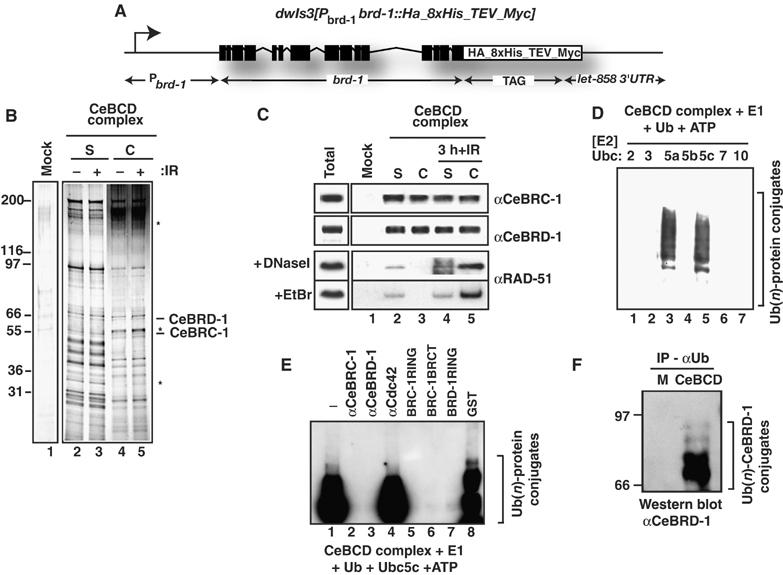
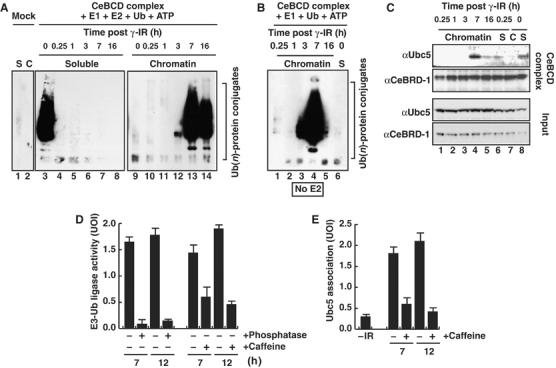
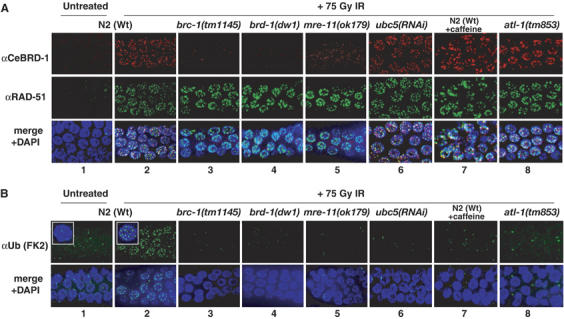
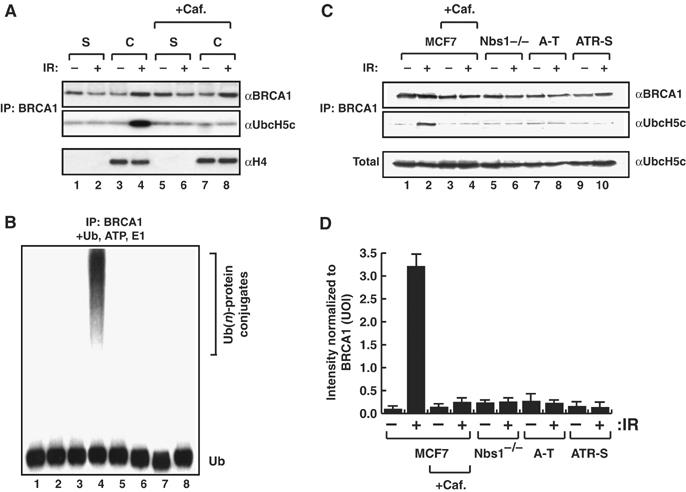
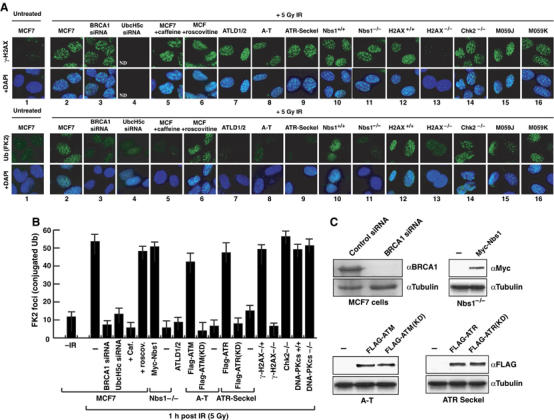
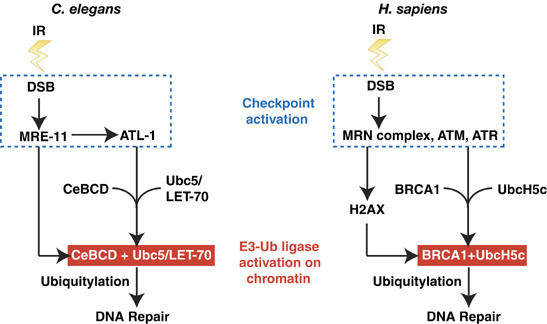
The BRCA1 tumour suppressor and its heterodimeric partner BARD1 constitute an E3-ubiquitin (Ub) ligase and function in DNA repair by unknown mechanisms. We show here that the Caenorhabditis elegans BRCA1/BARD1 (CeBCD) complex possesses an E3-Ub ligase responsible for ubiquitylation at DNA damage sites following ionizing radiation (IR). The DNA damage checkpoint promotes the association of the CeBCD complex with E2-Ub conjugating enzyme, Ubc5(LET-70), leading to the formation of an active E3-Ub ligase on chromatin following IR. Correspondingly, defects in Ubc5(let-70) or the DNA damage checkpoint genes atl-1 or mre-11 abolish CeBCD-dependent ubiquitylation in vivo. Extending these findings to human cells reveals a requirement for UbcH5c, the MRN complex, gamma-H2AX and a co-dependence for ATM and ATR kinases for BRCA1-dependent ubiquitylation at DNA damage sites. Furthermore, we demonstrate that the DNA damage checkpoint promotes the association between BRCA1 and UbcH5c to form an active E3-Ub ligase on chromatin after IR. These data reveal that BRCA1-dependent ubiquitylation is activated at sites of DNA repair by the checkpoint as part of a conserved DNA damage response.
Figures
References
-
- Alpi A, Pasierbek P, Gartner A, Loidl J (2003) Genetic and cytological characterization of the recombination protein RAD-51 in Caenorhabditis elegans. Chromosoma 112: 6–16 - PubMed
-
- Boulton SJ, Gartner A, Reboul J, Vaglio P, Dyson N, Hill DE, Vidal M (2002) Combined functional genomic maps of the C. elegans DNA damage response. Science 295: 127–131 - PubMed
-
- Boulton SJ, Martin JS, Polanowska J, Hill DE, Gartner A, Vidal M (2004) BRCA1/BARD1 orthologs required for DNA repair in Caenorhabditis elegans. Curr Biol 14: 33–39 - PubMed
-
- Bradbury JM, Jackson SP (2003) ATM and ATR. Curr Biol 13: R468. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Research Materials
Miscellaneous
