

Opaque cells signal white cells to form biofilms in Candida albicans
- PMID: 16628217
- PMCID: PMC1462973
- DOI: 10.1038/sj.emboj.7601099
Opaque cells signal white cells to form biofilms in Candida albicans
Abstract
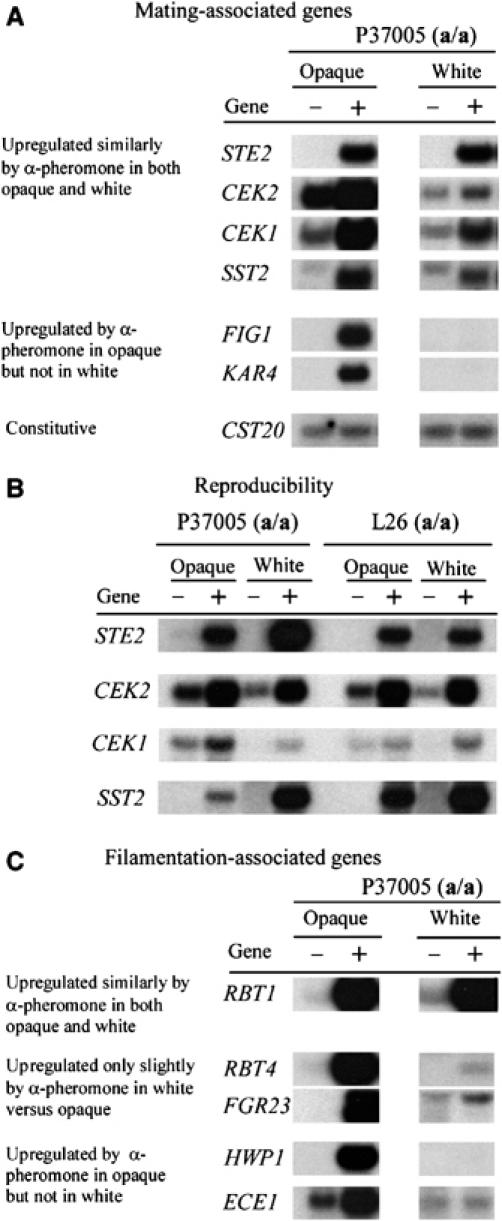
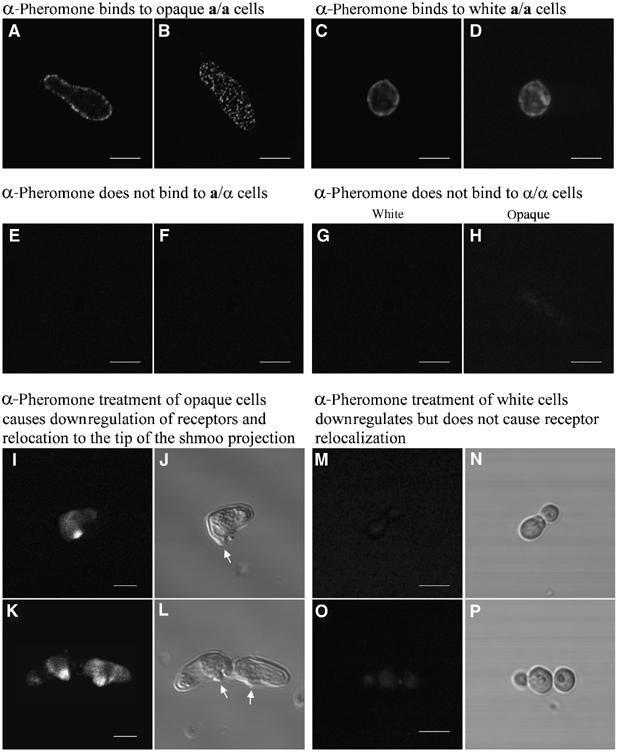
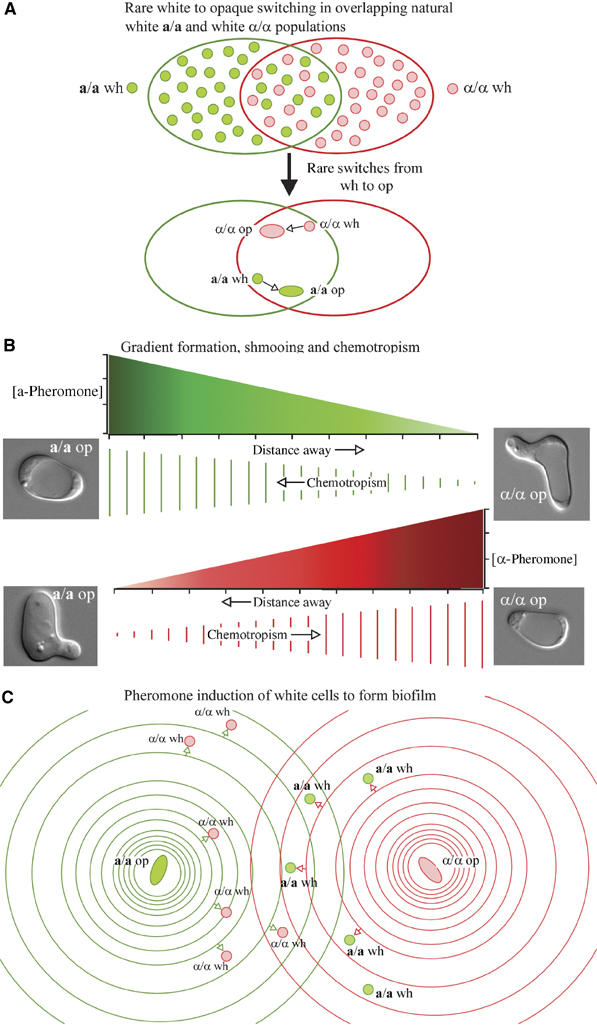
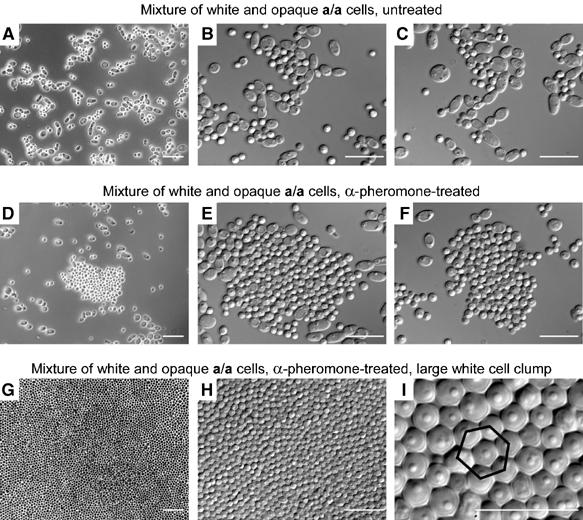
Upon homozygosis from a/alpha to a/a or alpha/alpha, Candida albicans must still switch from the 'white' to 'opaque' phenotype to mate. It was, therefore, surprising to discover that pheromone selectively upregulated mating-associated genes in mating-incompetent white cells without causing G1 arrest or shmoo formation. White cells, like opaque cells, possess pheromone receptors, although their distribution and redistribution upon pheromone treatment differ between the two cell types. In speculating about the possible role of the white cell pheromone response, it is hypothesized that in overlapping white a/a and alpha/alpha populations in nature, rare opaque cells, through the release of pheromone, signal majority white cells of opposite mating type to form a biofilm that facilitates mating. In support of this hypothesis, it is demonstrated that pheromone induces cohesiveness between white cells, minority opaque cells increase two-fold the thickness of majority white cell biofilms, and majority white cell biofilms facilitate minority opaque cell chemotropism. These results reveal a novel form of communication between switch phenotypes, analogous to the inductive events during embryogenesis in higher eukaryotes.
Figures

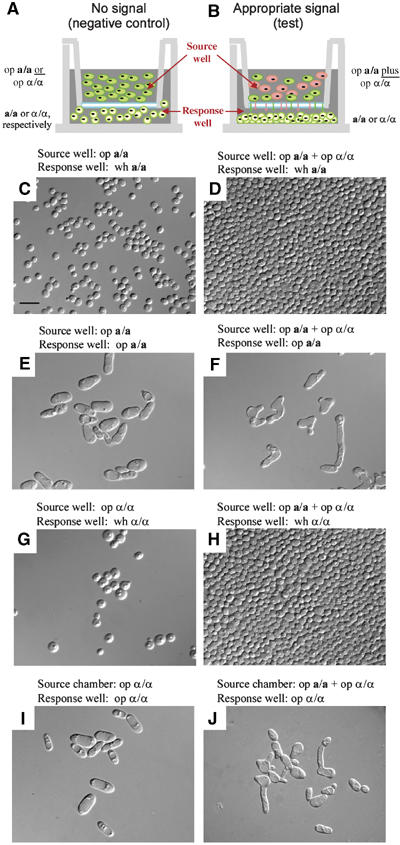
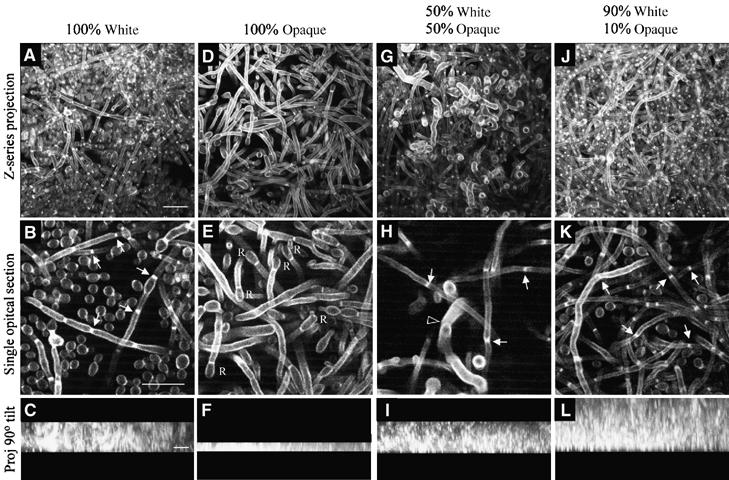
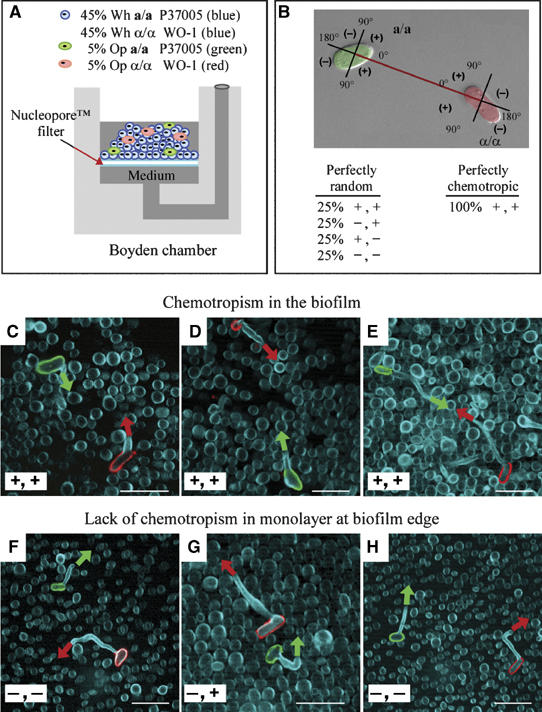
References
-
- Baillie GS, Douglas LJ (1999) Role of dimorphism in the development of Candida albicans biofilms. J Med Microbiol 48: 671–679 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
