

Interaction of Moloney murine leukemia virus matrix protein with IQGAP
- PMID: 16628219
- PMCID: PMC1462987
- DOI: 10.1038/sj.emboj.7601097
Interaction of Moloney murine leukemia virus matrix protein with IQGAP
Abstract
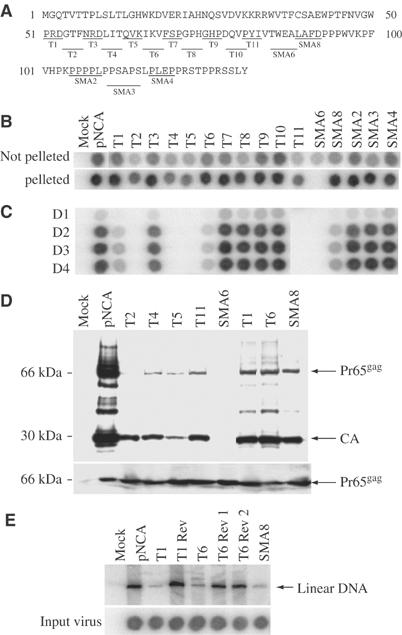
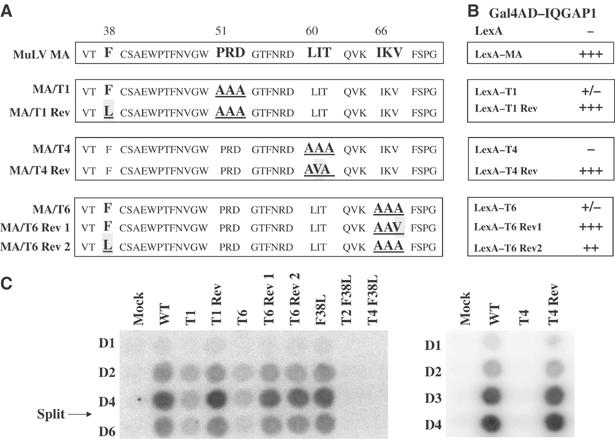
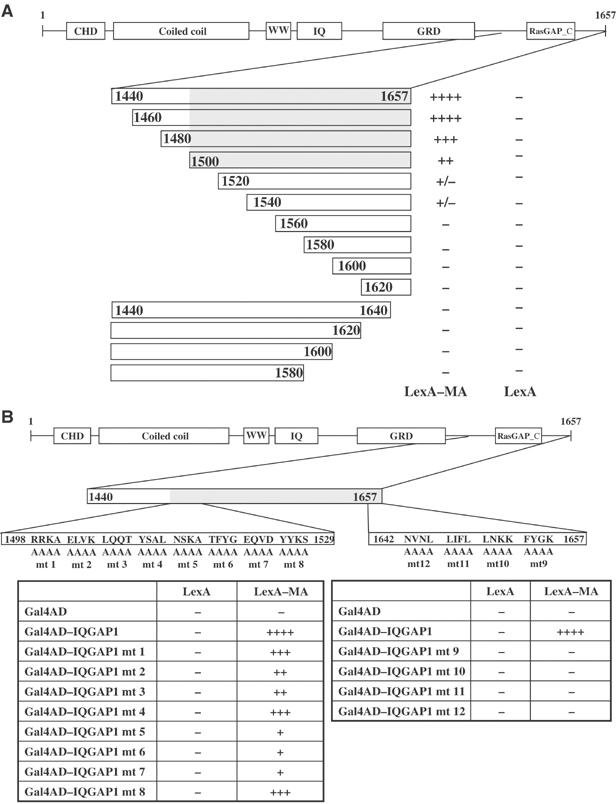
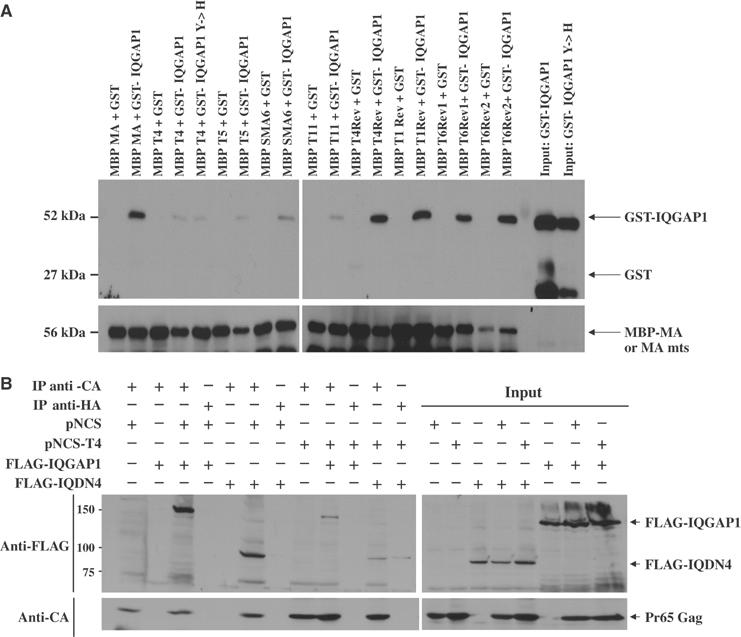
The matrix protein (MA) of the Moloney murine leukemia virus (M-MuLV) was found to interact with IQGAP1, a prominent regulator of the cytoskeleton. Mutational studies defined residues of MA critical for the interaction, and tests of viruses carrying MA mutations revealed a near-perfect correlation between binding and virus replication. The replication-defective mutants showed defects in both early and late stages of the life cycle. Four viable second-site revertant viruses were isolated from three different replication-defective parental mutants, and in all cases the interaction with IQGAP1 was restored by the suppressor mutations. The interaction of MA and IQGAP1 was readily detected in vitro and in vivo. Virus replication was potently inhibited by a C-terminal fragment of IQGAP1, and impaired by RNAi knockdown of IQGAP1 and 2. We suggest that the IQGAPs link the virus to the cytoskeleton for trafficking both into and out of the cell.
Figures
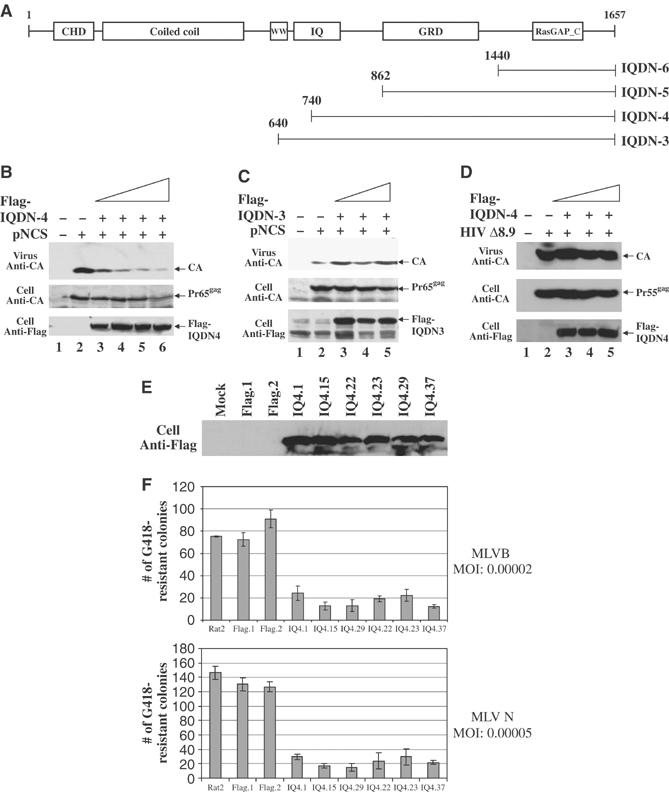
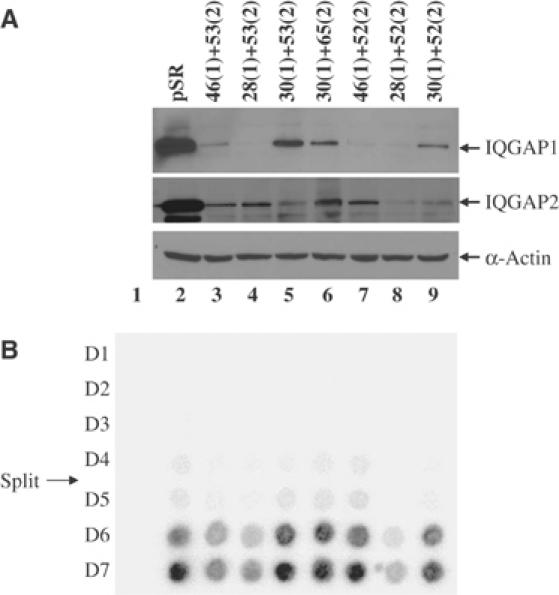
References
-
- Abramoff MD, Magelhaes PJ, Ram SJ (2004) Image processing with Image J. Biophoton Int 11: 36–42
-
- Basyuk E, Galli T, Mougel M, Blanchard JM, Sitbon M, Bertrand E (2003) Retroviral genomic RNAs are transported to the plasma membrane by endosomal vesicles. Dev Cell 5: 161–174 - PubMed
-
- Breedon L, Nasmyth K (1985) Regulation of the HO gene. Cold Spring Harbor Symp Quant Biol 50: 643–650 - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
