

pH-dependent conformational switch activates the inhibitor of transcription elongation
- PMID: 16628221
- PMCID: PMC1462974
- DOI: 10.1038/sj.emboj.7601094
pH-dependent conformational switch activates the inhibitor of transcription elongation
Abstract
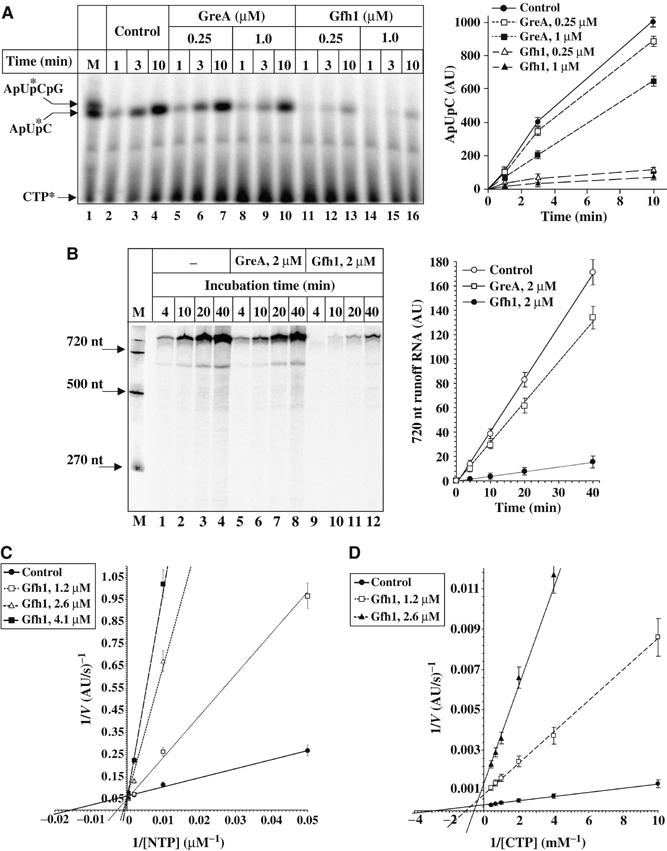
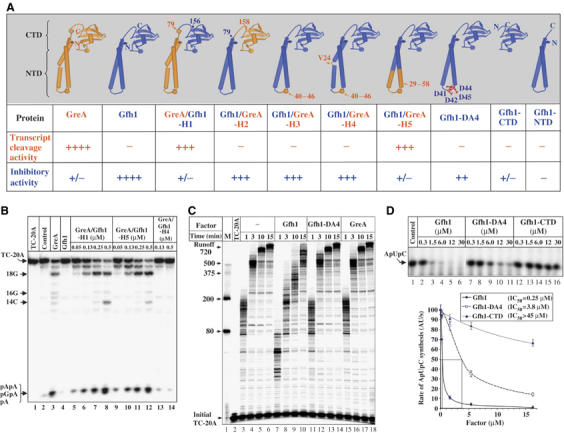
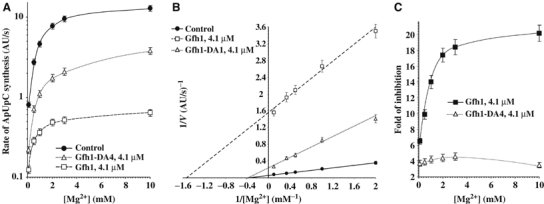
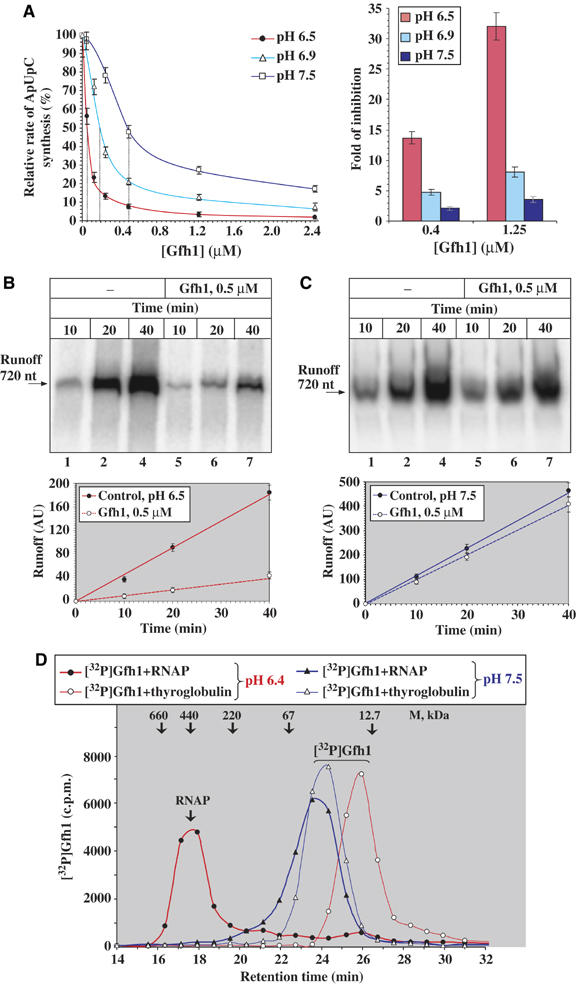
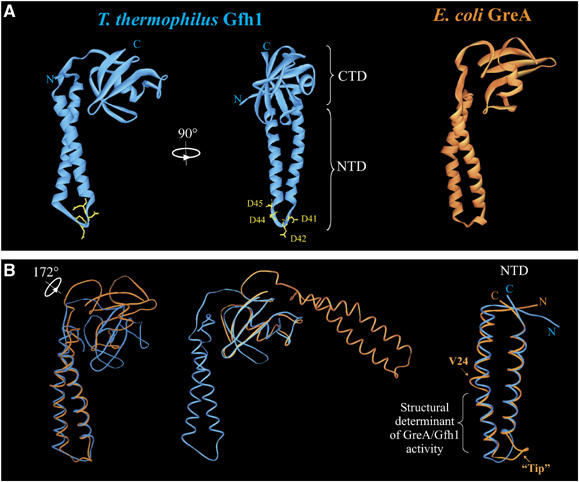
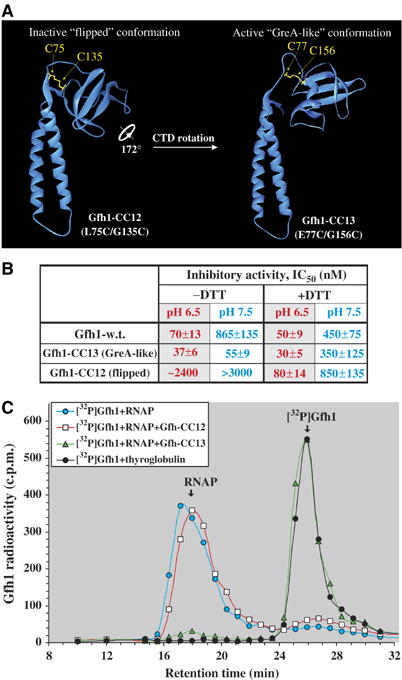
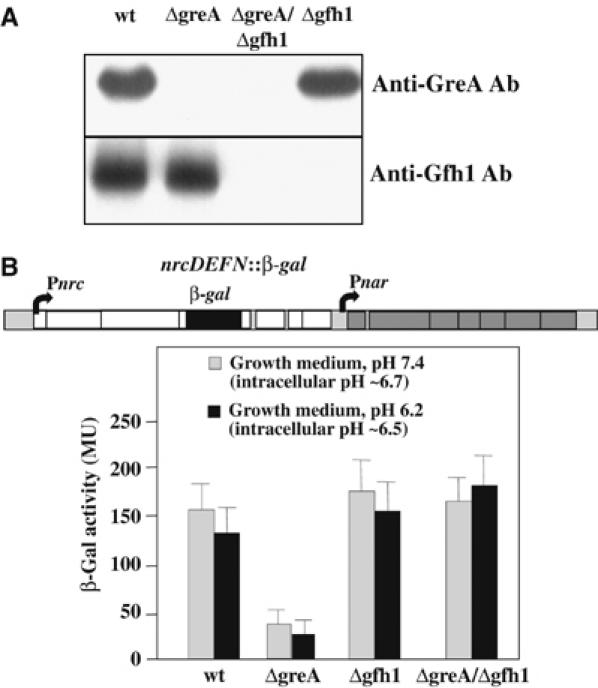
Gfh1, a transcription factor from Thermus thermophilus, inhibits all catalytic activities of RNA polymerase (RNAP). We characterized the Gfh1 structure, function and possible mechanism of action and regulation. Gfh1 inhibits RNAP by competing with NTPs for coordinating the active site Mg2+ ion. This coordination requires at least two aspartates at the tip of the Gfh1 N-terminal coiled-coil domain (NTD). The overall structure of Gfh1 is similar to that of the Escherichia coli transcript cleavage factor GreA, except for the flipped orientation of the C-terminal domain (CTD). We show that depending on pH, Gfh1-CTD exists in two alternative orientations. At pH above 7, it assumes an inactive 'flipped' orientation seen in the structure, which prevents Gfh1 from binding to RNAP. At lower pH, Gfh1-CTD switches to an active 'Gre-like' orientation, which enables Gfh1 to bind to and inhibit RNAP.
Figures
References
-
- Adelman K, Yuzenkova J, La Porta A, Zenkin N, Lee J, Lis JT, Borukhov S, Wang MD, Severinov K (2004) Molecular mechanism of transcription inhibition by peptide antibiotic Microcin J25. Mol Cell 14: 753–762 - PubMed
-
- Borukhov S, Lee J, Laptenko O (2005) Bacterial transcription elongation factors: new insights into molecular mechanism of action. Mol Microbiol 55: 1315–1324 - PubMed
-
- Brunger AT, Adams PD, Clore GM, DeLano WL, Gros P, Grosse-Kunstleve RW, Jiang JS, Kuszewski J, Nilges M, Pannu NS, Read RJ, Rice LM, Simonson T, Warren GL (1998) Crystallography & NMR system: a new software suite for macromolecular structure determination. Acta Crystallogr D 54: 905–921 - PubMed
-
- Cava F, de Pedro MA, Schwarz H, Henne A, Berenguer J (2004a) Binding to pyruvylated compounds as an ancestral mechanism to anchor the outer envelope in primitive bacteria. Mol Microbiol 52: 677–690 - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
