

Dorsal cochlear nucleus response properties following acoustic trauma: response maps and spontaneous activity
- PMID: 16630701
- PMCID: PMC1582886
- DOI: 10.1016/j.heares.2006.03.011
Dorsal cochlear nucleus response properties following acoustic trauma: response maps and spontaneous activity
Abstract
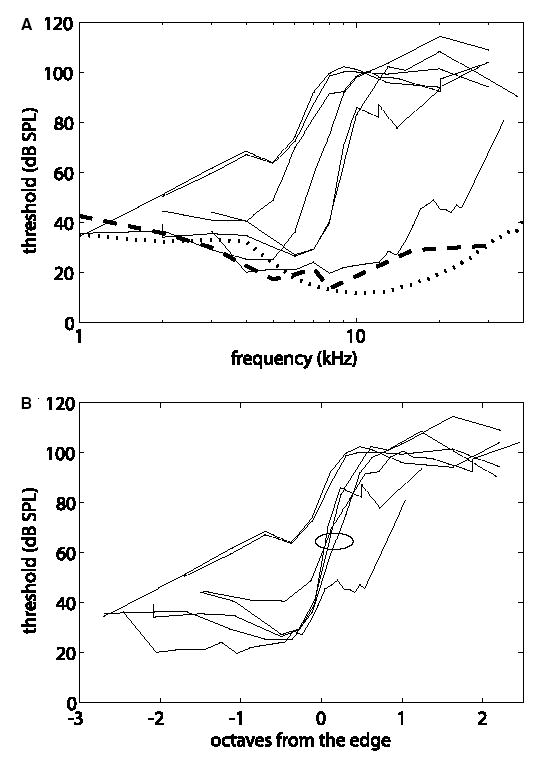
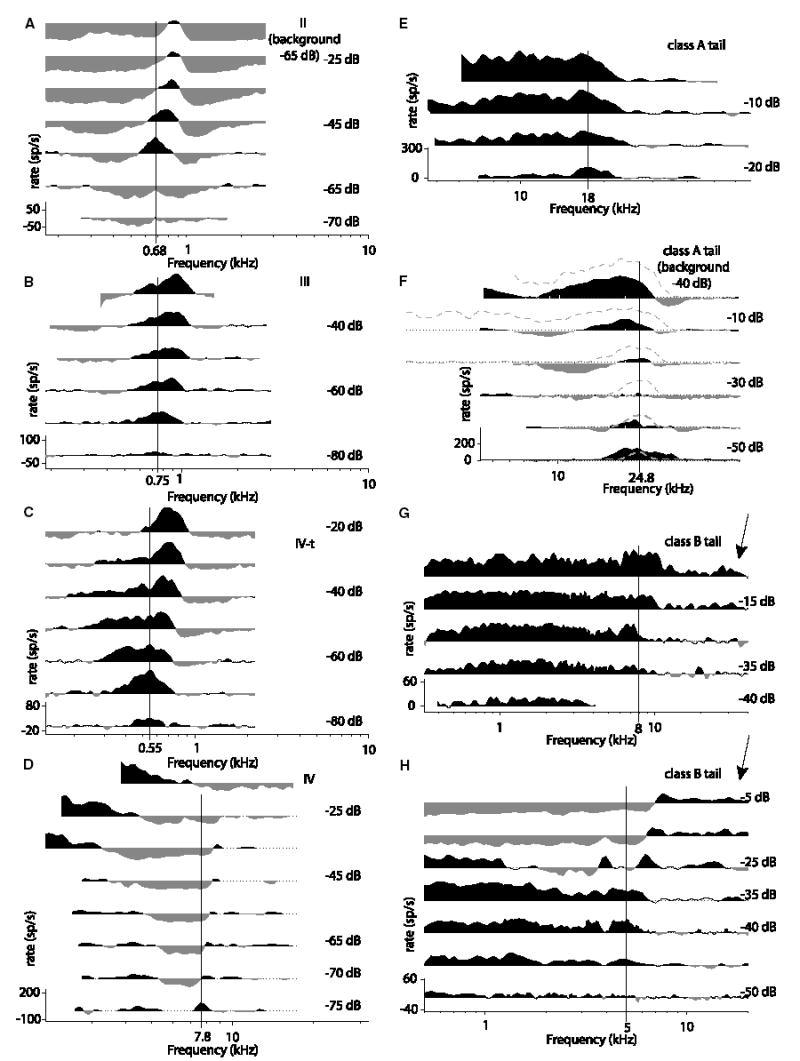
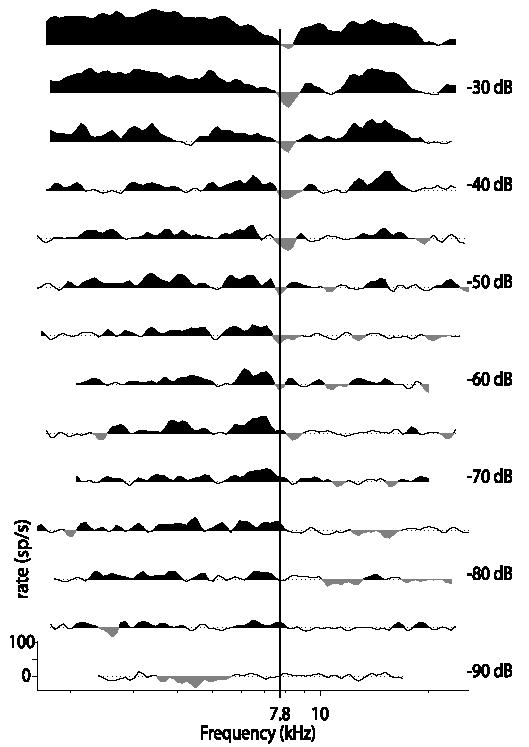
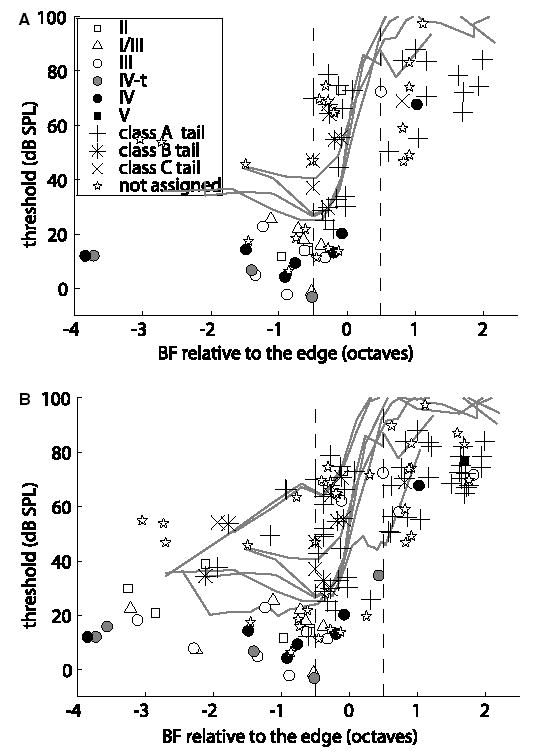
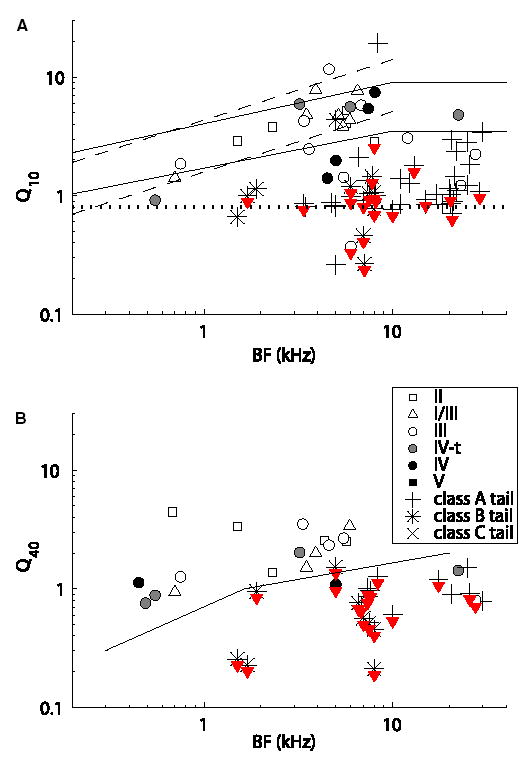
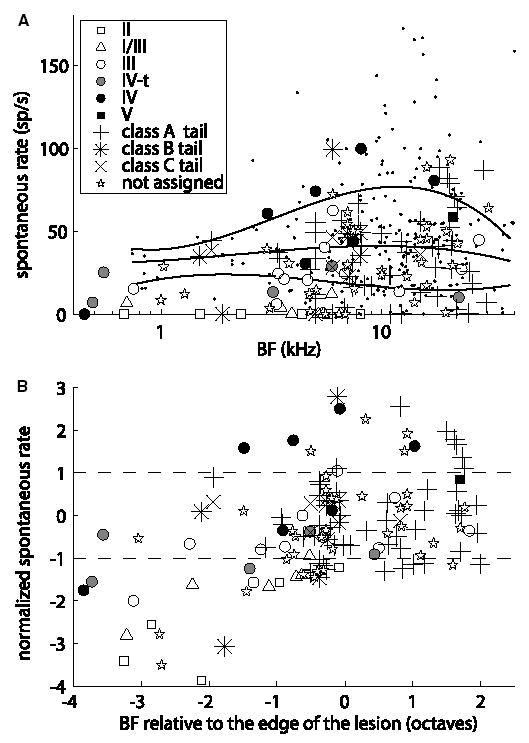
Recordings from single neurons in the dorsal cochlear nucleus (DCN) of unanesthetized (decerebrate) cats were done to characterize the effects of acoustic trauma. Trauma was produced by a 250 Hz band of noise centered at 10 kHz, presented at 105-120 dB SPL for 4h. After a one-month recovery period, neurons were recorded in the DCN. The threshold shift, determined from compound action-potential audiograms, showed a sharp threshold elevation of about 60 dB at BFs above an edge frequency of 5-10 kHz. The response maps of neurons with best frequencies (BFs) above the edge did not show the typical organization of excitatory and inhibitory areas seen in the DCN of unexposed animals. Instead, neurons showed no response to sound, weak responses that were hard to tune and characterize, or "tail" responses, consisting of broadly-tuned, predominantly excitatory responses, with a roughly low-pass shape similar to the tuning curves of auditory nerve fibers with similar threshold shifts. In some tail responses whose BFs were near the edge of the threshold elevation, a second weak high-frequency response was seen that suggests convergence of auditory nerve inputs with widely separated BFs on these cells. Spontaneous rates among neurons with elevated thresholds were not increased over those in populations of principal neurons in unexposed animals.
Figures
References
-
- Abbott SD, Hughes LF, Bauer CA, Salvi R, Caspary DM. Detection of glutamate decarboxylase isoforms in rat inferior colliculus following acoustic exposure. Neuroscience. 1999;93:1375–1381. - PubMed
-
- Anderson MJ, Young ED. Isoflurane/N2O anesthesia suppresses narrowband but not wideband inhibition in dorsal cochlear nucleus. Hear Res. 2004;188:29–41. - PubMed
-
- Berrebi AS, Mugnaini E. Distribution and targets of the cartwheel cell axon in the dorsal cochlear nucleus of the guinea pig. Anat Embryol. 1991;183:427–454. - PubMed
-
- Bilak M, Kim J, Potashner SJ, Bohne BA, Morest DK. New growth of axons in the cochlear nucleus of adult chinchillas after acoustic trauma. Exp Neurol. 1997;147:256–268. - PubMed
-
- Bledsoe SC, Jr, Nagase S, Miller JM, Altschuler RA. Deafness-induced plasticity in the mature central auditory system. Neuroreport. 1995;7:225–229. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Miscellaneous
