

In-frame deletion in a novel centrosomal/ciliary protein CEP290/NPHP6 perturbs its interaction with RPGR and results in early-onset retinal degeneration in the rd16 mouse
- PMID: 16632484
- PMCID: PMC1592550
- DOI: 10.1093/hmg/ddl107
In-frame deletion in a novel centrosomal/ciliary protein CEP290/NPHP6 perturbs its interaction with RPGR and results in early-onset retinal degeneration in the rd16 mouse
Abstract
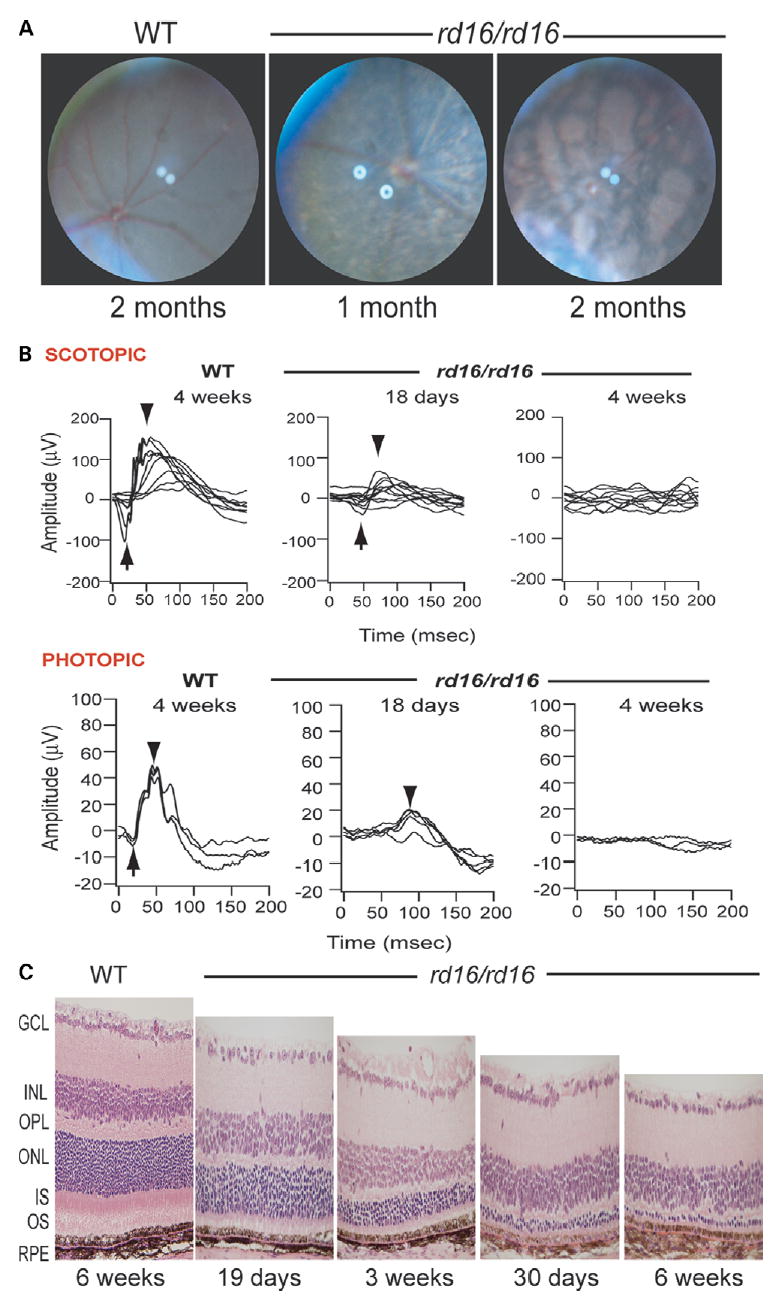
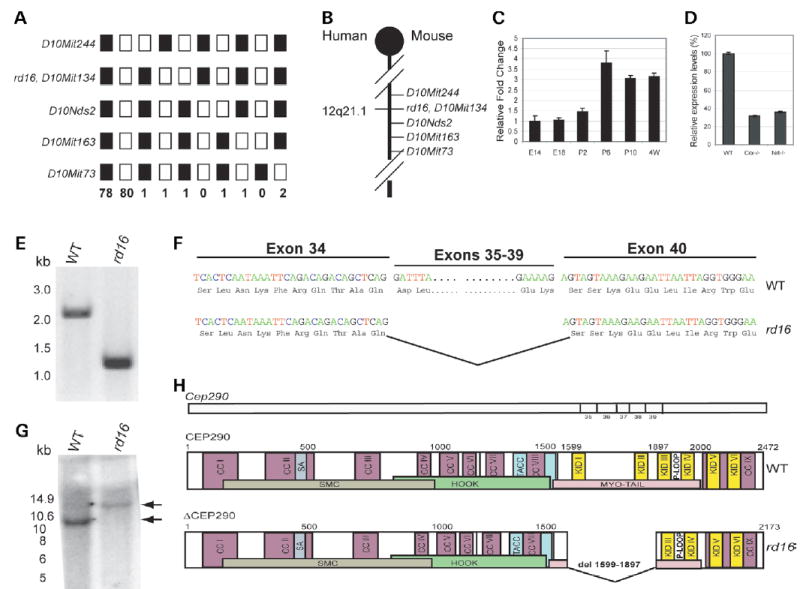
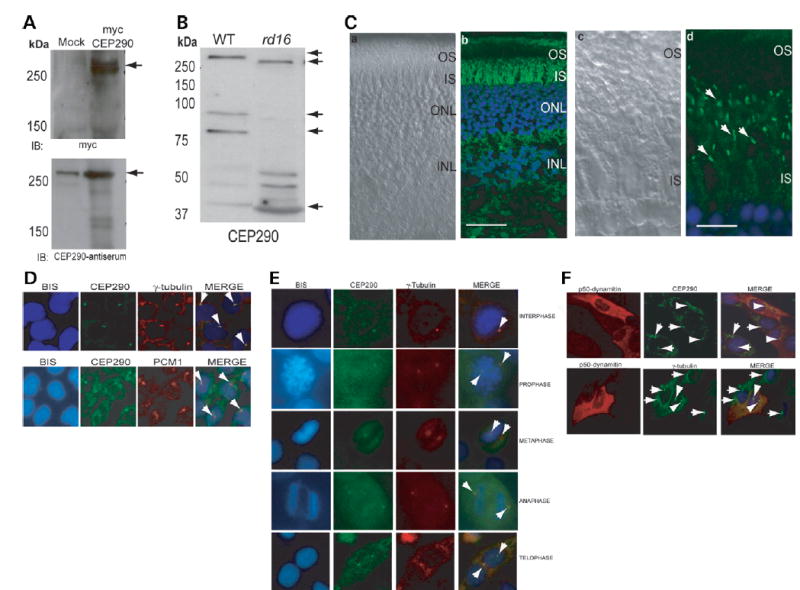
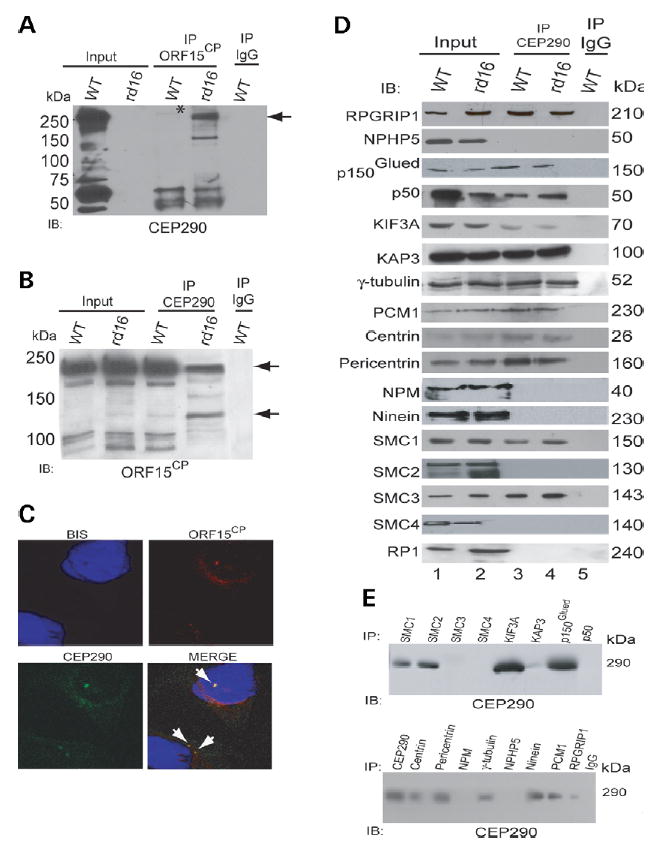
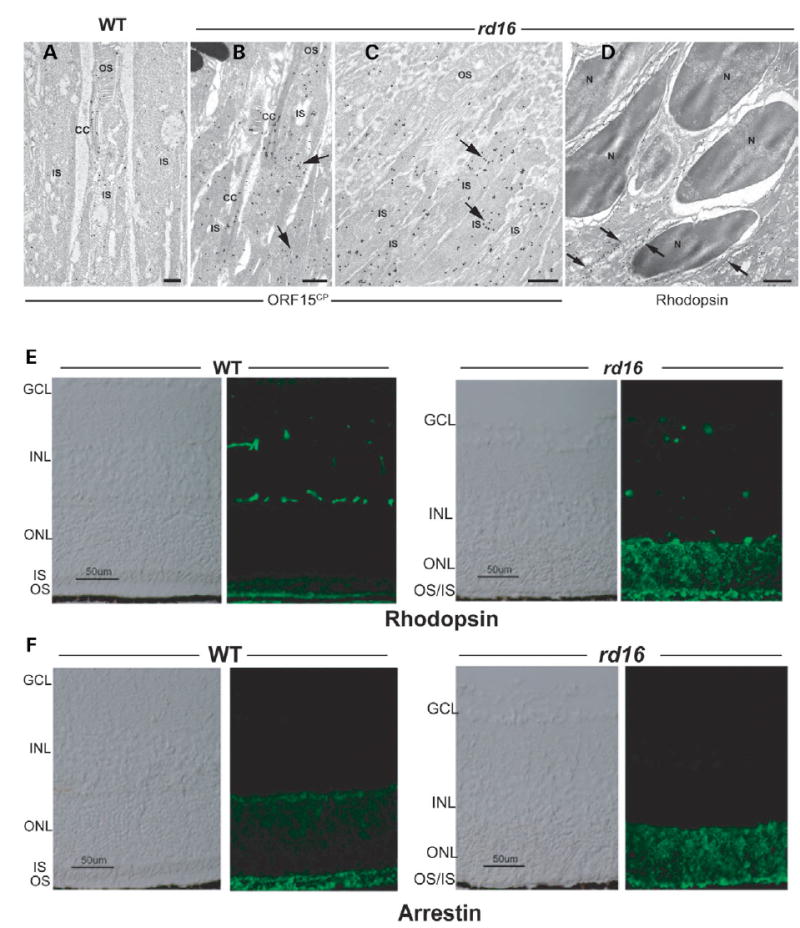
Centrosome- and cilia-associated proteins play crucial roles in establishing polarity and regulating intracellular transport in post-mitotic cells. Using genetic mapping and positional candidate strategy, we have identified an in-frame deletion in a novel centrosomal protein CEP290 (also called NPHP6), leading to early-onset retinal degeneration in a newly identified mouse mutant, rd16. We demonstrate that CEP290 localizes primarily to centrosomes of dividing cells and to the connecting cilium of retinal photoreceptors. We show that, in the retina, CEP290 associates with several microtubule-based transport proteins including RPGR, which is mutated in approximately 15% of patients with retinitis pigmentosa. A truncated CEP290 protein (DeltaCEP290) is detected in the rd16 retina, but in considerably reduced amounts; however, the mutant protein exhibits stronger association with specific RPGR isoform(s). Immunogold labeling studies demonstrate the redistribution of RPGR and of phototransduction proteins in the photoreceptors of rd16 retina. Our findings suggest a critical function for CEP290 in ciliary transport and provide insights into the mechanism of early-onset photoreceptor degeneration.
Conflict of interest statement
Figures
References
-
- Nigg EA. Centrosomes in Development and Disease. Wiley-VCH Verlag GmbH & Co; KGaA: 2004.
-
- Doxsey S. Re-evaluating centrosome function. Nat Rev Mol Cell Biol. 2001;2:688–698. - PubMed
-
- Beisson J, Wright M. Basal body/centriole assembly and continuity. Curr Opin Cell Biol. 2003;15:96–104. - PubMed
-
- Badano JL, Teslovich TM, Katsanis N. The centrosome in human genetic disease. Nat Rev Genet. 2005;6:194–205. - PubMed
-
- Young RW. Passage of newly formed protein through the connecting cilium of retina rods in the frog. J Ultrastruct Res. 1968;23:462–473. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
Grants and funding
- DK 1069274/DK/NIDDK NIH HHS/United States
- EY 12598/EY/NEI NIH HHS/United States
- DK 20572/DK/NIDDK NIH HHS/United States
- R01 EY007042/EY/NEI NIH HHS/United States
- P30 EY012598/EY/NEI NIH HHS/United States
- RR 01183/RR/NCRR NIH HHS/United States
- P60 DK020572/DK/NIDDK NIH HHS/United States
- R01 EY007758/EY/NEI NIH HHS/United States
- P30 EY007003/EY/NEI NIH HHS/United States
- F31 EY007003/EY/NEI NIH HHS/United States
- R01 EY013408/EY/NEI NIH HHS/United States
- EY 07961/EY/NEI NIH HHS/United States
- DK 10683-06/DK/NIDDK NIH HHS/United States
- P40 RR001183/RR/NCRR NIH HHS/United States
- R01 EY007961/EY/NEI NIH HHS/United States
- EY 07003/EY/NEI NIH HHS/United States
- P30 DK020572/DK/NIDDK NIH HHS/United States
- EY 13408/EY/NEI NIH HHS/United States
- DK 064614/DK/NIDDK NIH HHS/United States
- R01 DK064614/DK/NIDDK NIH HHS/United States
- EY 07758/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
