

Binding of 5'-GTP to the C-terminal FeS cluster of the radical S-adenosylmethionine enzyme MoaA provides insights into its mechanism
- PMID: 16632608
- PMCID: PMC1458979
- DOI: 10.1073/pnas.0510711103
Binding of 5'-GTP to the C-terminal FeS cluster of the radical S-adenosylmethionine enzyme MoaA provides insights into its mechanism
Abstract
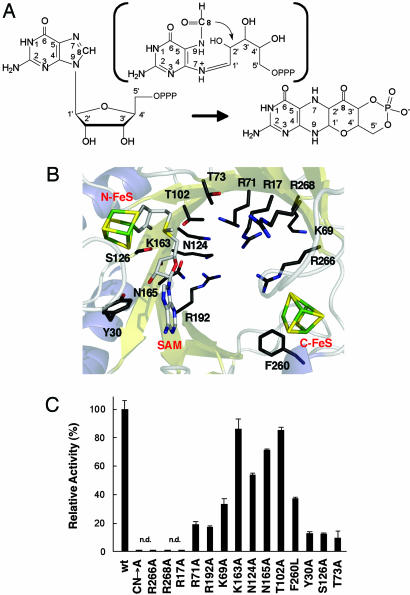
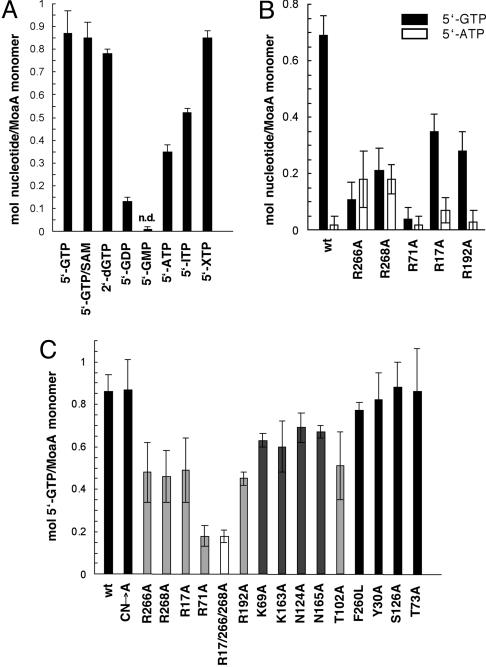
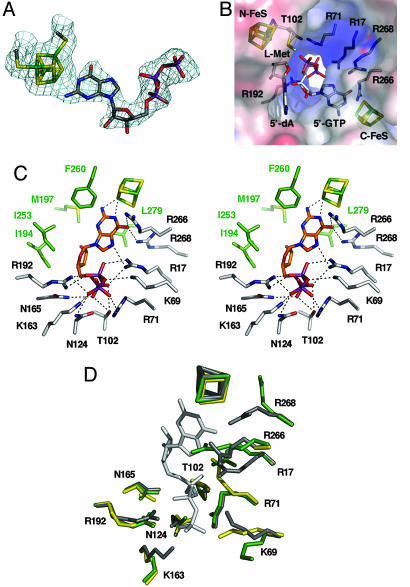
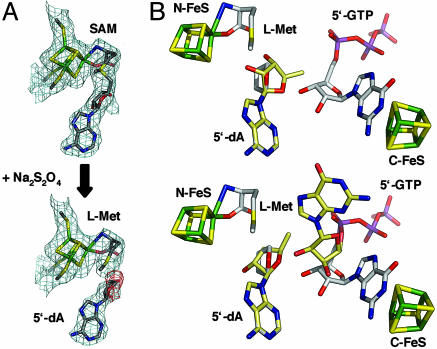
The first step in molybdenum cofactor biosynthesis, the conversion of 5'-GTP to precursor Z, an oxygen-sensitive tetrahydropyranopterin is catalyzed by the S-adenosylmethionine (SAM)-dependent enzyme MoaA and the accessory protein MoaC. This reaction involves the radical-initiated intramolecular rearrangement of the guanine C8 atom. MoaA harbors an N-terminal [4Fe-4S] cluster, which is involved in the reductive cleavage of SAM and generates a 5'-deoxyadenosyl radical (5'-dA*), and a C-terminal [4Fe-4S] cluster presumably involved in substrate binding and/or activation. Biochemical studies identified residues involved in 5'-GTP binding and the determinants of nucleotide specificity. The crystal structure of MoaA in complex with 5'-GTP confirms the biochemical data and provides valuable insights into the subsequent radical reaction. MoaA binds 5'-GTP with high affinity and interacts through its C-terminal [4Fe-4S] cluster with the guanine N1 and N2 atoms, in a yet uncharacterized binding mode. The tightly anchored triphosphate moiety prevents the escape of radical intermediates. This structure also visualizes the L-Met and 5'-dA cleavage products of SAM. Rotation of the 5'-dA ribose and/or conformational changes of the guanosine are proposed to bring the 5'-deoxyadenosyl radical into close proximity of either the ribose C2' and C3' or the guanine C8 carbon atoms leading to hydrogen abstraction.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Johnson D. C., Dean D. R., Smith A. D., Johnson M. K. Annu. Rev. Biochem. 2005;74:247–281. - PubMed
-
- Johnson M. K. Curr. Opin. Chem. Biol. 1998;2:173–181. - PubMed
-
- Beinert H., Holm R. H., Munck E. Science. 1997;277:653–659. - PubMed
-
- Beinert H. J. Biol. Inorg. Chem. 2000;5:2–15. - PubMed
-
- Flint D. H., Allen R. M. Chem. Rev. 1996;96:2315–2334. - PubMed
Publication types
MeSH terms
Substances
Associated data
- Actions
- Actions
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
