

The light peak of the electroretinogram is dependent on voltage-gated calcium channels and antagonized by bestrophin (best-1)
- PMID: 16636205
- PMCID: PMC2151522
- DOI: 10.1085/jgp.200509473
The light peak of the electroretinogram is dependent on voltage-gated calcium channels and antagonized by bestrophin (best-1)
Abstract
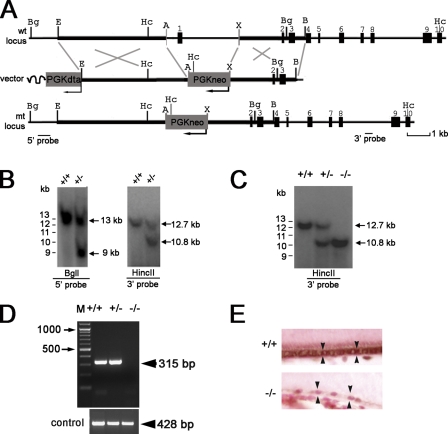
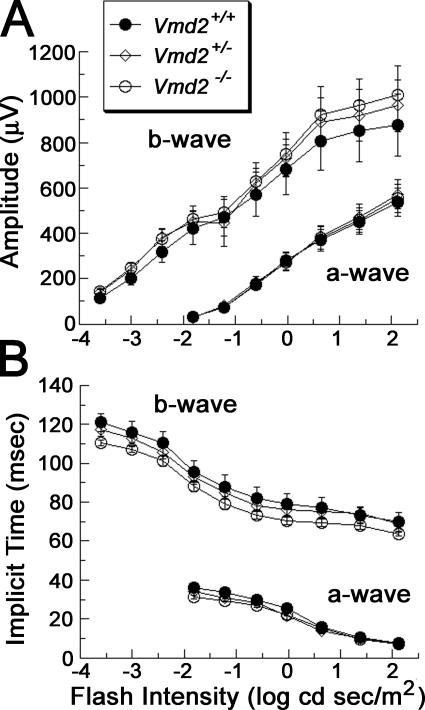
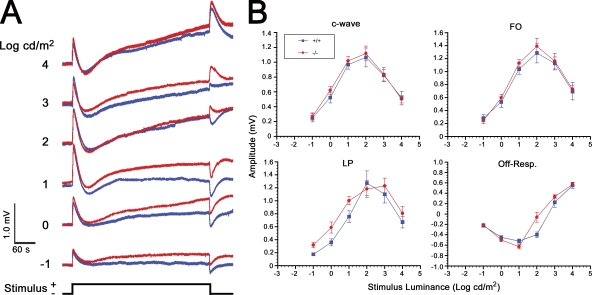
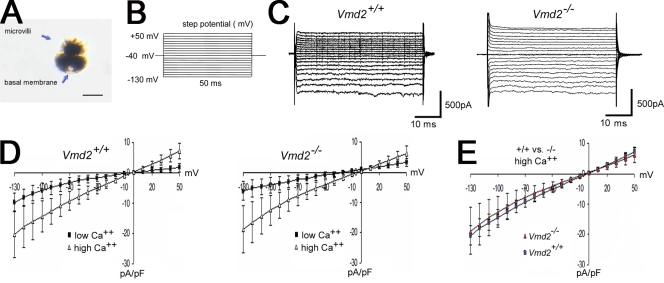
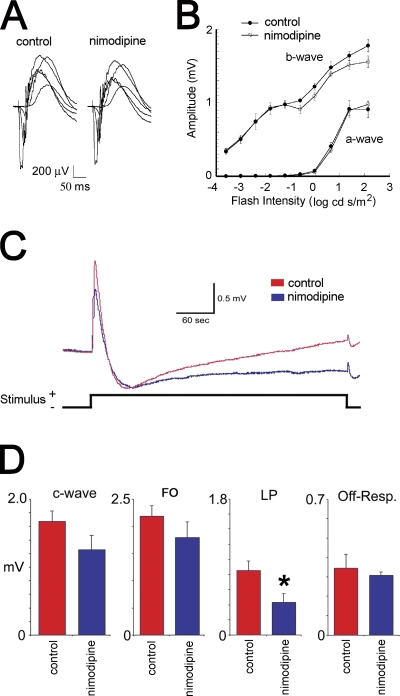
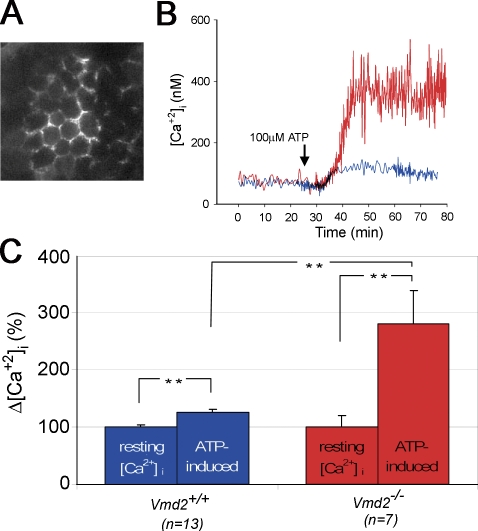
Mutations in VMD2, encoding bestrophin (best-1), cause Best vitelliform macular dystrophy (BMD), adult-onset vitelliform macular dystrophy (AVMD), and autosomal dominant vitreoretinochoroidopathy (ADVIRC). BMD is distinguished from AVMD by a diminished electrooculogram light peak (LP) in the absence of changes in the flash electroretinogram. Although the LP is thought to be generated by best-1, we find enhanced LP luminance responsiveness with normal amplitude in Vmd2-/- mice and no differences in cellular Cl- currents in comparison to Vmd2+/+ littermates. The putative Ca2+ sensitivity of best-1, and our recent observation that best-1 alters the kinetics of voltage-dependent Ca2+ channels (VDCC), led us to examine the role of VDCCs in the LP. Nimodipine diminished the LP, leading us to survey VDCC beta-subunit mutant mice. Lethargic mice, which harbor a loss of function mutation in the beta4 subunit of VDCCs, exhibited a significant shift in LP luminance response, establishing a role for Ca2+ in LP generation. When stimulated with ATP, which increases [Ca++]I, retinal pigment epithelial cells derived from Vmd2-/- mice exhibited a fivefold greater response than Vmd2+/+ littermates, indicating that best-1 can suppress the rise in [Ca2+]I associated with the LP. We conclude that VDCCs regulated by a beta4 subunit are required to generate the LP and that best-1 antagonizes the LP luminance response potentially via its ability to modulate VDCC function. Furthermore, we suggest that the loss of vision associated with BMD is not caused by the same pathologic process as the diminished LP, but rather is caused by as yet unidentified effects of best-1 on other cellular processes.
Figures


References
-
- Allikmets, R., J.M. Seddon, P.S. Bernstein, A. Hutchinson, A. Atkinson, S. Sharma, B. Gerrard, W. Li, M.L. Metzker, C. Wadelius, et al. 1999. Evaluation of the Best disease gene in patients with age-related macular degeneration and other maculopathies. Hum. Genet. 104:449–453. - PubMed
-
- Bakall, B., L.Y. Marmorstein, G. Hoppe, N.S. Peachey, C. Wadelius, and A.D. Marmorstein. 2003. Expression and localization of bestrophin during normal mouse development. Invest. Ophthalmol. Vis. Sci. 44:3622–3628. - PubMed
-
- Ball, S.L., P.A. Powers, H.-S. Shin, C.W. Morgans, N.S. Peachey, and R.G. Gregg. 2002. Role of the β2 subunit of voltage-dependent calcium channels in the retinal outer plexiform layer. Invest. Ophthalmol. Vis. Sci. 43:1595–1603. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
- R01 EY012354/EY/NEI NIH HHS/United States
- EY14465/EY/NEI NIH HHS/United States
- EY12354/EY/NEI NIH HHS/United States
- EY15638/EY/NEI NIH HHS/United States
- R01 EY014465/EY/NEI NIH HHS/United States
- EY14898/EY/NEI NIH HHS/United States
- EY13160/EY/NEI NIH HHS/United States
- EY13847/EY/NEI NIH HHS/United States
- R24 EY015638/EY/NEI NIH HHS/United States
- R01 EY013847/EY/NEI NIH HHS/United States
- R56 EY013160/EY/NEI NIH HHS/United States
- R01 EY013160/EY/NEI NIH HHS/United States
- R03 EY014898/EY/NEI NIH HHS/United States
LinkOut - more resources
Full Text Sources
Other Literature Sources
Molecular Biology Databases
Miscellaneous
