

Disulfide cross-linked protein represents a significant fraction of ALS-associated Cu, Zn-superoxide dismutase aggregates in spinal cords of model mice
- PMID: 16636274
- PMCID: PMC1447524
- DOI: 10.1073/pnas.0602048103
Disulfide cross-linked protein represents a significant fraction of ALS-associated Cu, Zn-superoxide dismutase aggregates in spinal cords of model mice
Abstract
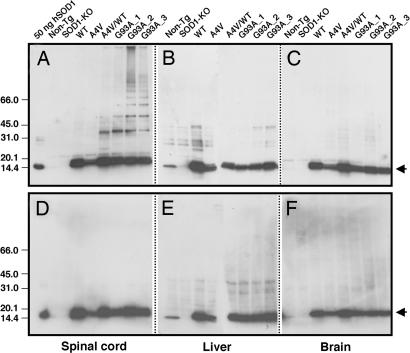
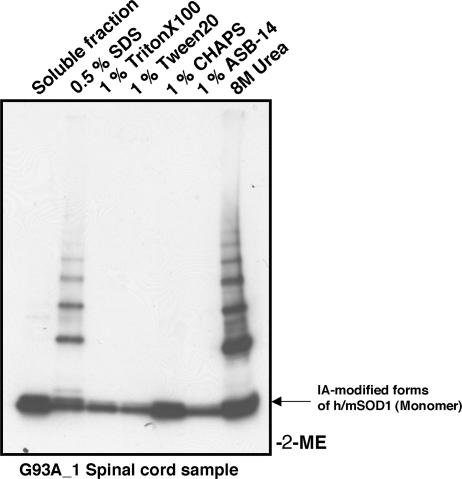
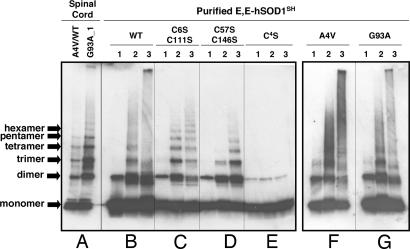
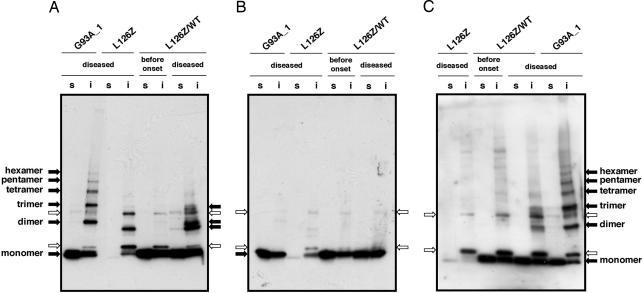
Point mutations in Cu, Zn-superoxide dismutase (SOD1) cause a familial form of the neurodegenerative disease amyotrophic lateral sclerosis (ALS). Aggregates of mutant SOD1 proteins are observed in histopathology and are invoked in several proposed mechanisms for motor neuronal death; however, the significant stability and activity of the mature mutant proteins are not readily explained in such models. Recent biochemical studies suggest that it is the immature disulfide-reduced forms of the familial ALS mutant SOD1 proteins that play a critical role; these forms tend to misfold, oligomerize, and readily undergo incorrect disulfide formation upon mild oxidative stress in vitro. Here we provide physiological support for this mechanism of aggregate formation and show that a significant fraction of the insoluble SOD1 aggregates in spinal cord of the ALS-model transgenic mice contain multimers cross-linked via intermolecular disulfide bonds. These insoluble disulfide-linked SOD1 multimers are found only in the spinal cord of symptomatic transgenic animals, are not observed in unafflicted tissue such as brain cortex and liver, and can incorporate WT SOD1 protein. The findings provide a biochemical basis for a pathological hallmark of this disease; namely, incorrect disulfide cross-linking of the immature, misfolded mutant proteins leads to insoluble aggregates.
Conflict of interest statement
Conflict of interest statement: No conflicts declared.
Figures
References
-
- Rosen D. R., Siddique T., Patterson D., Figlewicz D. A., Sapp P., Hentati A., Donaldson D., Goto J., O'Regan J. P., Deng H. X., et al. Nature. 1993;362:59–62. - PubMed
-
- Deng H. X., Hentati A., Tainer J. A., Iqbal Z., Cayabyab A., Hung W. Y., Getzoff E. D., Hu P., Herzfeldt B., Roos R. P., et al. Science. 1993;261:1047–1051. - PubMed
-
- McCord J. M., Fridovich I. J. Biol. Chem. 1969;244:6049–6055. - PubMed
-
- Reaume A. G., Elliott J. L., Hoffman E. K., Kowall N. W., Ferrante R. J., Siwek D. F., Wilcox H. M., Flood D. G., Beal M. F., Brown R. H., Jr, et al. Nat. Genet. 1996;13:43–47. - PubMed
-
- Liochev S. I., Fridovich I. Free Radical Biol. Med. 2003;34:1383–1389. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Medical
Molecular Biology Databases
Miscellaneous
