

3-Nitrotyrosine as a spectroscopic probe for investigating protein protein interactions
- PMID: 16641485
- PMCID: PMC2242503
- DOI: 10.1110/ps.051957006
3-Nitrotyrosine as a spectroscopic probe for investigating protein protein interactions
Abstract
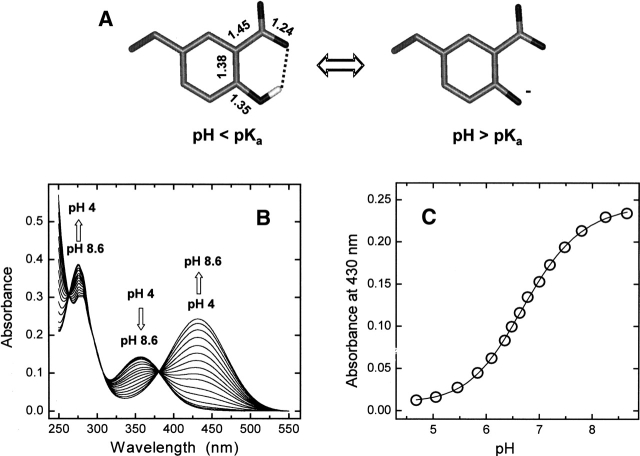
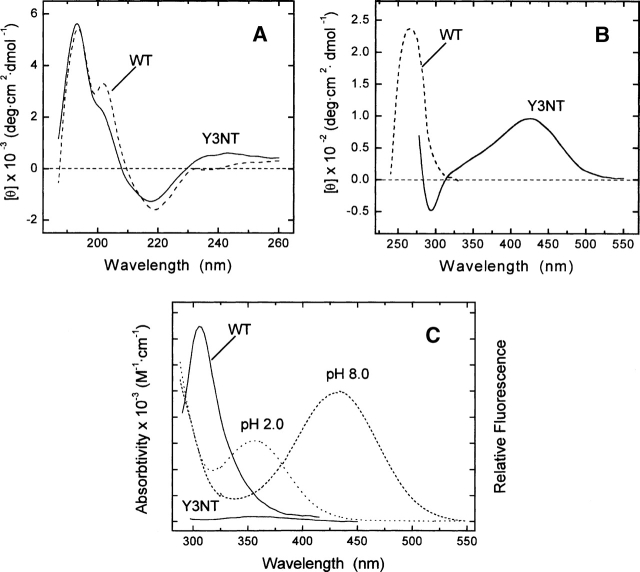
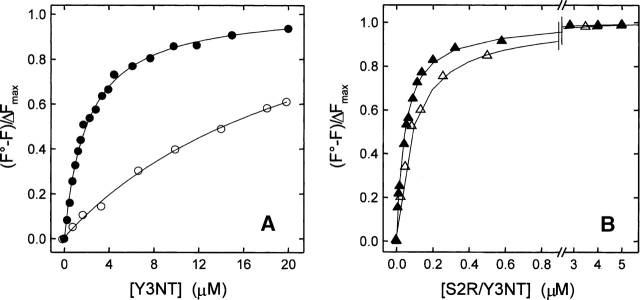
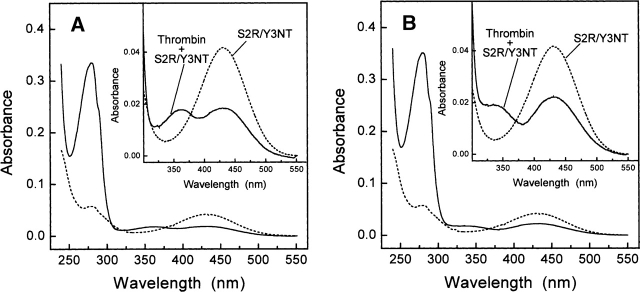
3-Nitrotyrosine (NT) is approximately 10(3)-fold more acidic than Tyr, and its absorption properties are strongly pH-dependent. NT absorbs radiation in the wavelength range where Tyr and Trp emit fluorescence (300-450 nm), and it is essentially nonfluorescent. Therefore, NT may function as an energy acceptor in resonance energy transfer (FRET) studies for investigating ligand protein interactions. Here, the potentialities of NT were tested on the hirudin thrombin system, a well-characterized protease inhibitor pair of key pharmacological importance. We synthesized two analogs of the N-terminal domain (residues 1-47) of hirudin: Y3NT, in which Tyr3 was replaced by NT, and S2R/Y3NT, containing the substitutions Ser2 --> Arg and Tyr3 --> NT. The binding of these analogs to thrombin was investigated at pH 8 by FRET and UV/Vis-absorption spectroscopy. Upon hirudin binding, the fluorescence of thrombin was reduced by approximately 50%, due to the energy transfer occurring between the Trp residues of the enzyme (i.e., the donors) and the single NT of the inhibitor (i.e., the acceptor). The changes in the absorption spectra of the enzyme inhibitor complex indicate that the phenate moiety of NT in the free state becomes protonated to phenol in the thrombin-bound form. Our results indicate that the incorporation of NT can be effectively used to detect protein protein interactions with sensitivity in the low nanomolar range, to uncover subtle structural features at the ligand protein interface, and to obtain reliable Kd values for structure activity relationship studies. Furthermore, advances in chemical and genetic methods, useful for incorporating noncoded amino acids into proteins, highlight the broad applicability of NT in biotechnology and pharmacological screening.
Figures


References
-
- Abraham M.H., Du C.M., Platts J.A. 2000. Lipophilicity of the nitrophenols J. Org. Chem. 65: 7114–7118. - PubMed
-
- Albericio F. 2004. Developments in peptides and amide synthesis Curr. Opin. Chem. Biol. 8: 211–221. - PubMed
-
- Bell R., Stevens W.K., Jia Z., Samis J., Coté H.C.F., MacGillivray R.T.A., Nesheim M.E. 2000. Fluorescence properties and functional roles of tryptophan residues 60d, 96, 148, and 215 of thrombin J. Biol. Chem. 275: 29513–29520. - PubMed
-
- Bode W., Turk D., Karshikov A. 1992. The refined 1.9-Å X-ray crystal structure of D-Phe-Pro-Arg chloromethylketone-inhibited human α-thrombin: Structure analysis, overall structure, electrostatic properties, detailed active-site geometry, and structure-function relationships Protein Sci. 1: 426–471. - PMC - PubMed
-
- Copeland R.A. In Enzymes: A practical introduction to structure, mechanism, and data analysis . 2000. 2d ed J. Wiley & Sons, New York.
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Other Literature Sources
Research Materials
