

Biosynthesis of wybutosine, a hyper-modified nucleoside in eukaryotic phenylalanine tRNA
- PMID: 16642040
- PMCID: PMC1462984
- DOI: 10.1038/sj.emboj.7601105
Biosynthesis of wybutosine, a hyper-modified nucleoside in eukaryotic phenylalanine tRNA
Abstract
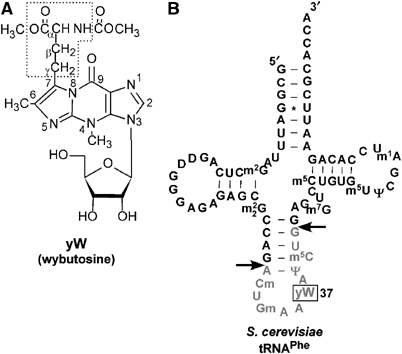
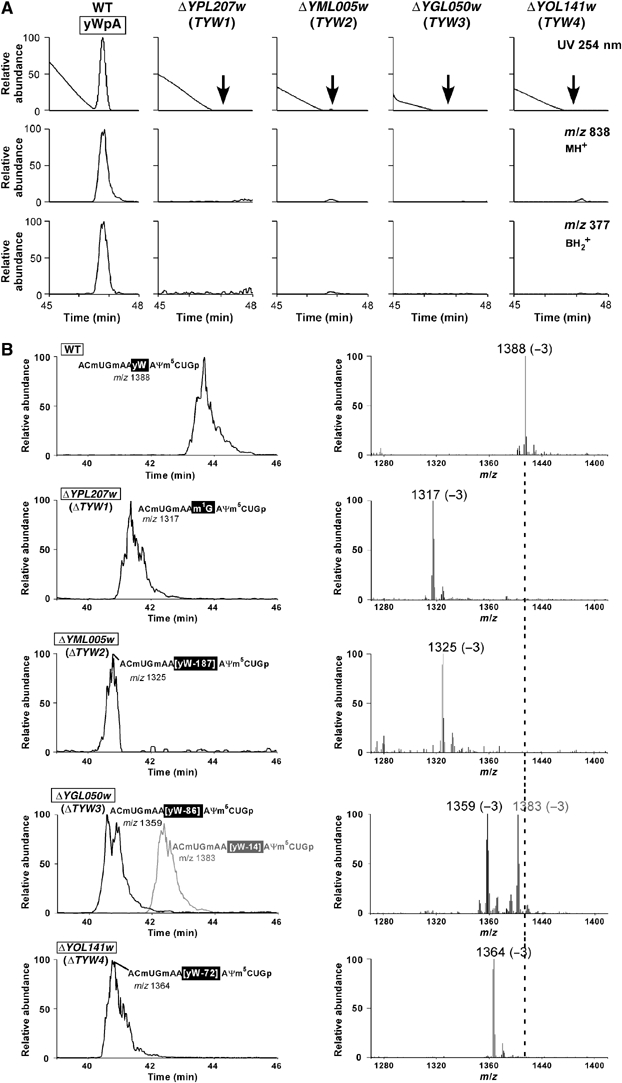
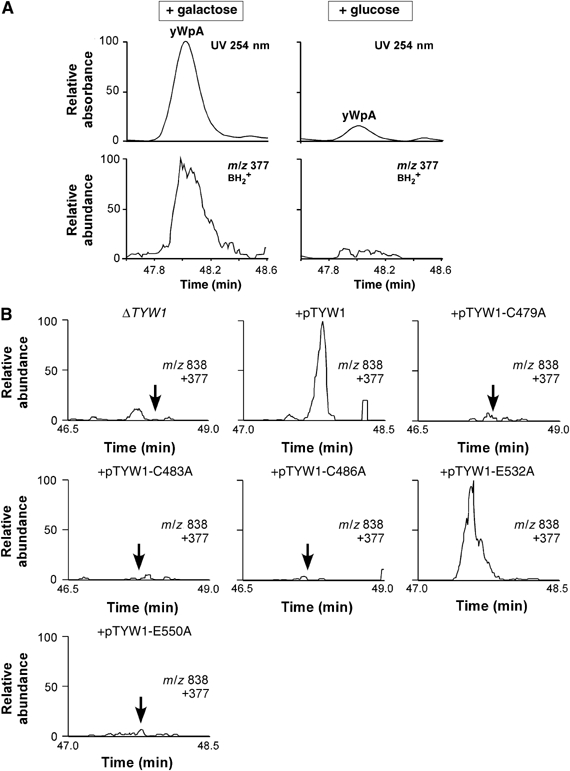
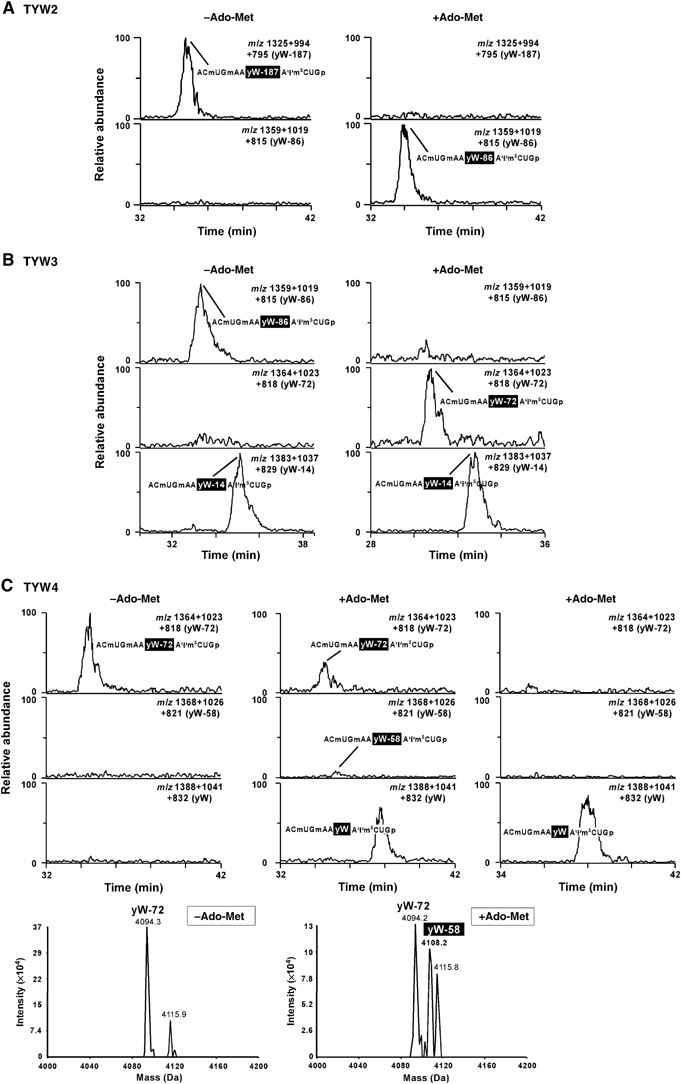
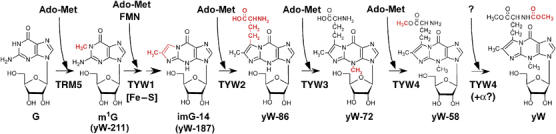
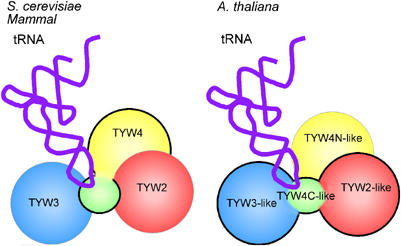
Wybutosine (yW) is a tricyclic nucleoside with a large side chain found at the 3'-position adjacent to the anticodon of eukaryotic phenylalanine tRNA. yW supports codon recognition by stabilizing codon-anticodon interactions during decoding on the ribosome. To identify genes responsible for yW synthesis from uncharacterized genes of Saccharomyces cerevisiae, we employed a systematic reverse genetic approach combined with mass spectrometry ('ribonucleome analysis'). Four genes YPL207w, YML005w, YGL050w and YOL141w (named TYW1, TYW2, TYW3 and TYW4, respectively) were essential for yW synthesis. Mass spectrometric analysis of each modification intermediate of yW revealed its sequential biosynthetic pathway. TYW1 is an iron-sulfur (Fe-S) cluster protein responsible for the tricyclic formation. Multistep enzymatic formation of yW from yW-187 could be reconstituted in vitro using recombinant TYW2, TYW3 and TYW4 with S-adenosylmethionine, suggesting that yW synthesis might proceed through sequential reactions in a complex formed by multiple components assembled with the precursor tRNA. This hypothesis is also supported by the fact that plant ortholog is a large fusion protein consisting of TYW2 and TYW3 with the C-terminal domain of TYW4.
Figures



Similar articles
-
Ribonucleome analysis identified enzyme genes responsible for wybutosine synthesis.Nucleic Acids Symp Ser (Oxf). 2006;(50):65-6. doi: 10.1093/nass/nrl032. Nucleic Acids Symp Ser (Oxf). 2006. PMID: 17150819
-
Structural basis of tRNA modification with CO2 fixation and methylation by wybutosine synthesizing enzyme TYW4.Nucleic Acids Res. 2009 May;37(9):2910-25. doi: 10.1093/nar/gkp158. Epub 2009 Mar 14. Nucleic Acids Res. 2009. PMID: 19287006 Free PMC article.
-
Structural basis of AdoMet-dependent aminocarboxypropyl transfer reaction catalyzed by tRNA-wybutosine synthesizing enzyme, TYW2.Proc Natl Acad Sci U S A. 2009 Sep 15;106(37):15616-21. doi: 10.1073/pnas.0905270106. Epub 2009 Aug 26. Proc Natl Acad Sci U S A. 2009. PMID: 19717466 Free PMC article.
-
Wybutosine biosynthesis: structural and mechanistic overview.RNA Biol. 2014;11(12):1508-18. doi: 10.4161/15476286.2014.992271. RNA Biol. 2014. PMID: 25629788 Free PMC article. Review.
-
Biosynthesis of wyosine derivatives in tRNA(Phe) of Archaea: role of a remarkable bifunctional tRNA(Phe):m1G/imG2 methyltransferase.RNA. 2014 Jun;20(6):747-53. doi: 10.1261/rna.043315.113. RNA. 2014. PMID: 24837075 Free PMC article. Review.
Cited by
-
Roles of Fe-S proteins: from cofactor synthesis to iron homeostasis to protein synthesis.Curr Opin Genet Dev. 2016 Jun;38:45-51. doi: 10.1016/j.gde.2016.03.006. Epub 2016 Apr 7. Curr Opin Genet Dev. 2016. PMID: 27061491 Free PMC article. Review.
-
Ribosome biogenesis factor Tsr3 is the aminocarboxypropyl transferase responsible for 18S rRNA hypermodification in yeast and humans.Nucleic Acids Res. 2016 May 19;44(9):4304-16. doi: 10.1093/nar/gkw244. Epub 2016 Apr 15. Nucleic Acids Res. 2016. PMID: 27084949 Free PMC article.
-
The ribose methylation enzyme FTSJ1 has a conserved role in neuron morphology and learning performance.Life Sci Alliance. 2023 Jan 31;6(4):e202201877. doi: 10.26508/lsa.202201877. Print 2023 Apr. Life Sci Alliance. 2023. PMID: 36720500 Free PMC article.
-
Site-Specific Profiling of 4-Thiouridine Across Transfer RNA Genes in Escherichia coli.ACS Omega. 2022 Jan 27;7(5):4011-4025. doi: 10.1021/acsomega.1c05071. eCollection 2022 Feb 8. ACS Omega. 2022. PMID: 35155896 Free PMC article.
-
Aquifex aeolicus tRNA (N2,N2-guanine)-dimethyltransferase (Trm1) catalyzes transfer of methyl groups not only to guanine 26 but also to guanine 27 in tRNA.J Biol Chem. 2009 Jul 31;284(31):20467-78. doi: 10.1074/jbc.M109.020024. Epub 2009 Jun 2. J Biol Chem. 2009. PMID: 19491098 Free PMC article.
References
-
- Björk GR (1995) Biosynthesis and function of modified nucleosides. In Söll D, Rajbhandary UL (eds). tRNA: Structure, Biosynthesis, and Function, pp 165–205. Washington, DC: ASM Press
-
- Blobstein SH, Grunberger D, Weinstein IB, Nakanishi K (1973) Isolation and structure determination of the fluorescent base from bovine liver phenylalanine transfer ribonucleic acid. Biochemistry 12: 188–193 - PubMed
-
- Brule H, Elliott M, Redlak M, Zehner ZE, Holmes WM (2004) Isolation and characterization of the human tRNA-(N1G37) methyltransferase (TRM5) and comparison to the Escherichia coli TrmD protein. Biochemistry 43: 9243–9255 - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
Miscellaneous
