

Regulation of rat hepatic L-pyruvate kinase promoter composition and activity by glucose, n-3 polyunsaturated fatty acids, and peroxisome proliferator-activated receptor-alpha agonist
- PMID: 16644726
- PMCID: PMC2766394
- DOI: 10.1074/jbc.M601277200
Regulation of rat hepatic L-pyruvate kinase promoter composition and activity by glucose, n-3 polyunsaturated fatty acids, and peroxisome proliferator-activated receptor-alpha agonist
Abstract
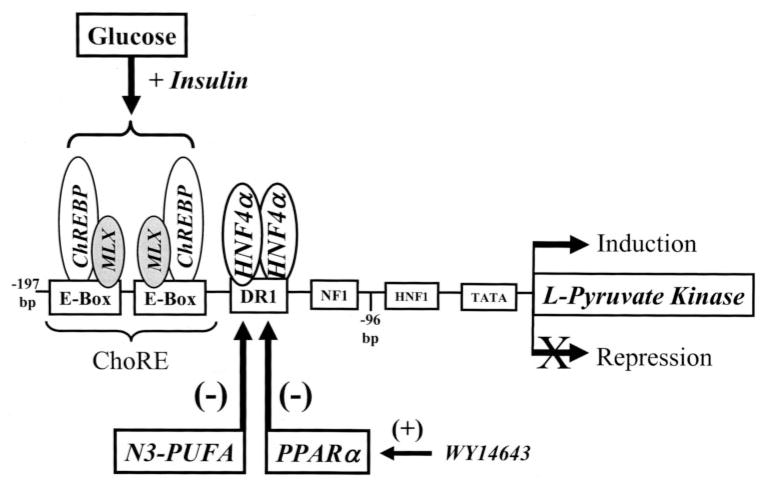
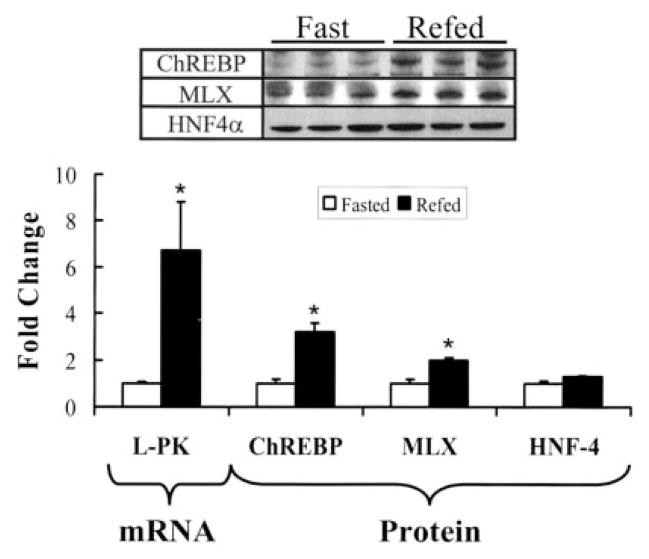
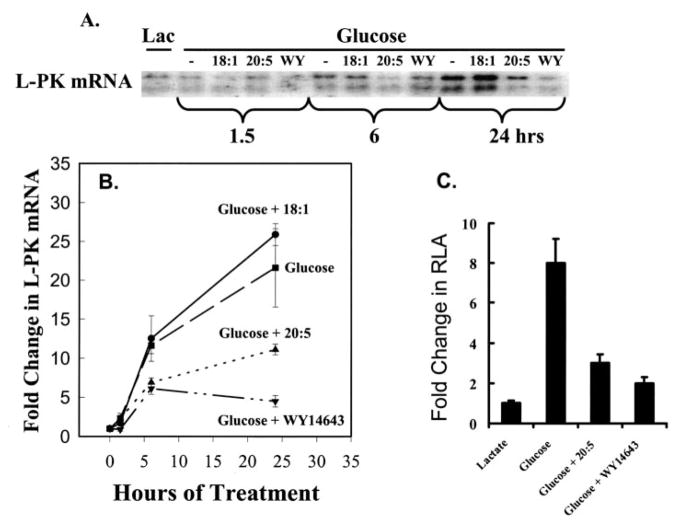
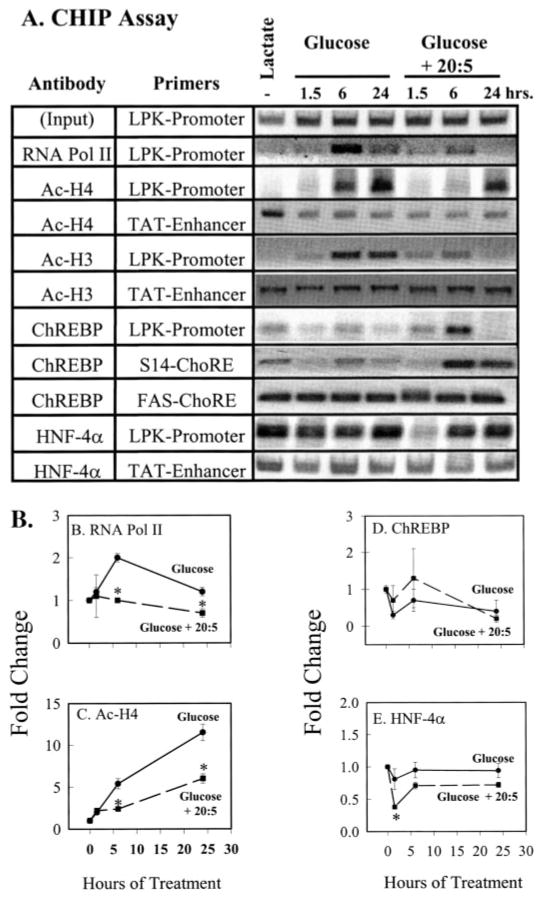
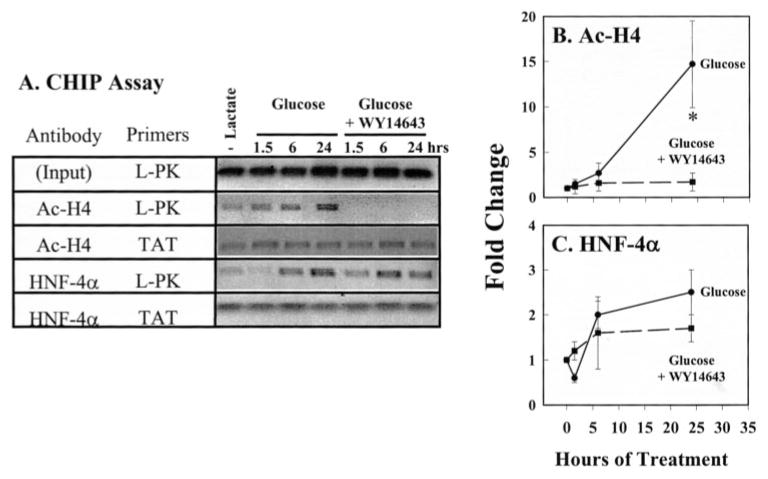
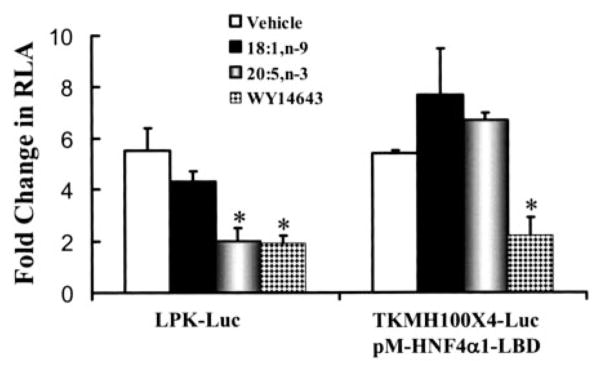
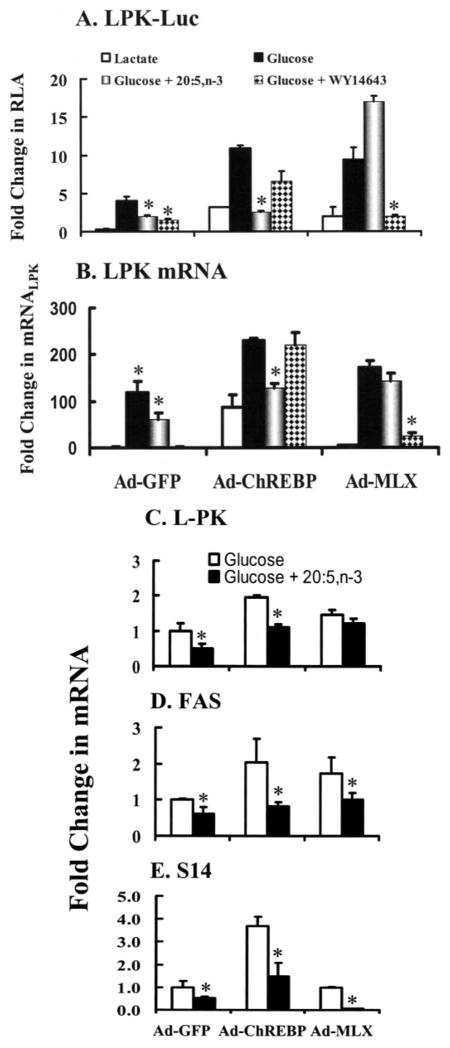
Carbohydrate regulatory element-binding protein (ChREBP), MAX-like factor X (MLX), and hepatic nuclear factor-4alpha (HNF-4alpha) are key transcription factors involved in the glucose-mediated induction of hepatic L-type pyruvate kinase (L-PK) gene transcription. n-3 polyunsaturated fatty acids (PUFA) and WY14643 (peroxisome proliferator-activated receptor alpha (PPARalpha) agonist) interfere with glucose-stimulated L-PK gene transcription in vivo and in rat primary hepatocytes. Feeding rats a diet containing n-3 PUFA or WY14643 suppressed hepatic mRNA(L-PK) but did not suppress hepatic ChREBP or HNF-4alpha nuclear abundance. Hepatic MLX nuclear abundance, however, was suppressed by n-3 PUFA but not WY14643. In rat primary hepatocytes, glucose-stimulated accumulation of mRNA(LPK) and L-PK promoter activity correlated with increased ChREBP nuclear abundance. This treatment also increased L-PK promoter occupancy by RNA polymerase II (RNA pol II), acetylated histone H3 (Ac-H3), and acetylated histone H4 (Ac-H4) but did not significantly impact L-PK promoter occupancy by ChREBP or HNF-4alpha. Inhibition of L-PK promoter activity by n-3 PUFA correlated with suppressed RNA pol II, Ac-H3, and Ac-H4 occupancy on the L-PK promoter. Although n-3 PUFA transiently suppressed ChREBP and MLX nuclear abundance, this treatment did not impact ChREBP-LPK promoter interaction. HNF4alpha-LPK promoter interaction was transiently suppressed by n-3 PUFA. Inhibition of L-PK promoter activity by WY14643 correlated with a transient decline in ChREBP nuclear abundance and decreased Ac-H4 interaction with the L-PK promoter. WY14643, however, had no impact on MLX nuclear abundance or HNF4alpha-LPK promoter interaction. Although overexpressed ChREBP or HNF-4alpha did not relieve n-3 PUFA suppression of L-PK gene expression, overexpressed MLX fully abrogated n-3 PUFA suppression of L-PK promoter activity and mRNA(L-PK) abundance. Overexpressed ChREBP, but not MLX, relieved the WY14643 inhibition of L-PK. In conclusion, n-3 PUFA and WY14643/PPARalpha target different transcription factors to control L-PK gene transcription. MLX, the heterodimer partner for ChREBP, has emerged as a novel target for n-3 PUFA regulation.
Figures


References
-
- Granner D, Pilkis S. J Biol Chem. 1990;265:10173–10176. - PubMed
-
- Jump DB, Botolin D, Wang Y, Xu J, Christian B, Demeure O. J Nutr. 2005;135:2503–2506. - PubMed
-
- Jump DB. CRC Crit Rev Clin Lab Sci. 2004;41:41–78. - PubMed
-
- Towle HC. Trends Endocrinol Metab. 2005;16:489–494. - PubMed
-
- Liimatta M, Towle HC, Clarke S, Jump DB. Mol Endocrinol. 1994;8:1147–1153. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
