

The cation-chloride cotransporter NKCC1 promotes sharp waves in the neonatal rat hippocampus
- PMID: 16644806
- PMCID: PMC1779742
- DOI: 10.1113/jphysiol.2006.107086
The cation-chloride cotransporter NKCC1 promotes sharp waves in the neonatal rat hippocampus
Abstract
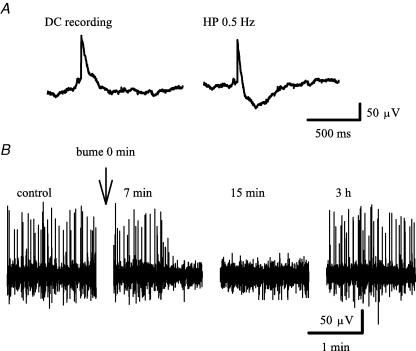
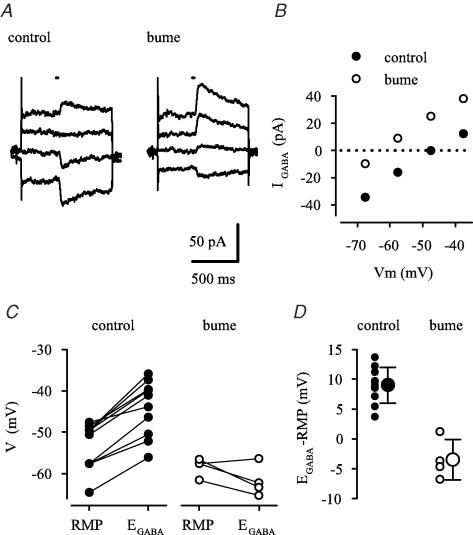
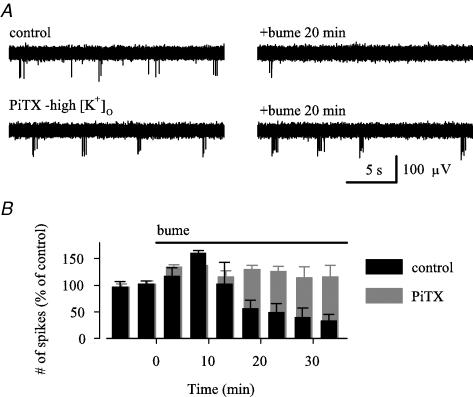
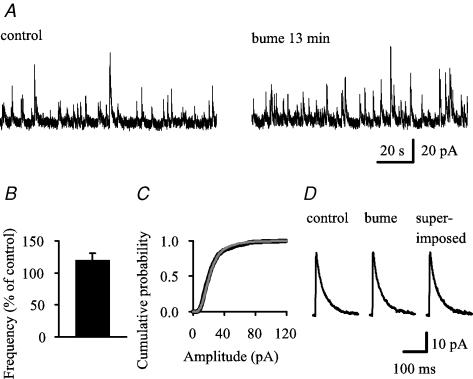
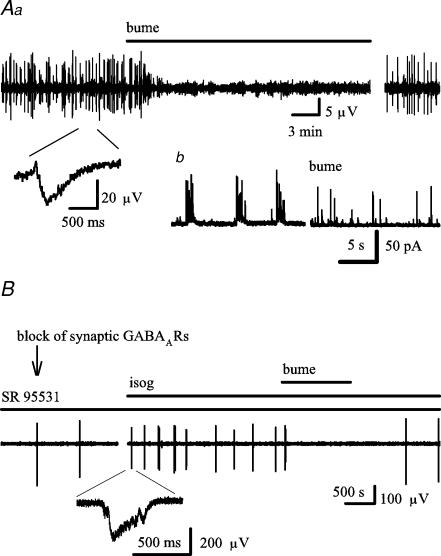
Earlier studies indicate a crucial role for the interconnected network of intrinsically bursting CA3 pyramidal neurons in the generation of in vivo hippocampal sharp waves (SPWs) and their proposed neonatal in vitro counterparts, the giant depolarizing potentials (GDPs). While mechanisms involving ligand- and voltage-gated channels have received lots of attention in the generation of CA3 network events in the immature hippocampus, the contribution of ion-transport mechanisms has not been extensively studied. Here, we show that bumetanide, a selective inhibitor of neuronal Cl- uptake mediated by the Na+-K+-2Cl- cotransporter isoform 1 (NKCC1), completely and reversibly blocks SPWs in the neonate (postnatal days 7-9) rat hippocampus in vivo, an action also seen on GDPs in slices (postnatal days 1-8). These findings strengthen the view that GDPs and early SPWs are homologous events. Gramicidin-perforated patch recordings indicated that NKCC1 accounts for a large ( approximately 10 mV) depolarizing driving force for the GABAA current in the immature CA3 pyramids. Consistent with a reduction in the depolarization mediated by endogenous GABAA-receptor activation, bumetanide inhibited the spontaneous bursts of individual neonatal CA3 pyramids, but it slightly increased the interneuronal activity as seen in the frequency of spontaneous GABAergic currents. An inhibitory effect of bumetanide was seen on the in vitro population events in the absence of synaptic GABAA receptor-mediated transmission, provided that a tonic GABAA receptor-mediated current was present. Our work indicates that NKCC1 expressed in CA3 pyramidal neurons promotes network activity in the developing hippocampus.
Figures
References
-
- Barry PH. JPCalc, a software package for calculating liquid junction potential corrections in patch-clamp, intracellular, epithelial and bilayer measurements and for correcting junction potential measurements. J Neurosci Meth. 1994;51:107–116. - PubMed
-
- Ben Ari Y. Developing networks play a similar melody. Trends Neurosci. 2001;24:353–360. - PubMed
-
- Bolea S, Avignone E, Berretta N, Sanchez-Andres JV, Cherubini E. Glutamate controls the induction of GABA-mediated giant depolarizing potentials through AMPA receptors in neonatal rat hippocampal slices. J Neurophysiol. 1999;81:2095–2102. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Miscellaneous