

Conditions leading to high CO2 (>5 kPa) in waterlogged-flooded soils and possible effects on root growth and metabolism
- PMID: 16644893
- PMCID: PMC3291891
- DOI: 10.1093/aob/mcl076
Conditions leading to high CO2 (>5 kPa) in waterlogged-flooded soils and possible effects on root growth and metabolism
Abstract
Aims: Soil waterlogging impedes gas exchange with the atmosphere, resulting in low P(O2) and often high P(CO2). Conditions conducive to development of high P(CO2) (5-70 kPa) during soil waterlogging and flooding are discussed. The scant information on responses of roots to high P(CO2) in terms of growth and metabolism is reviewed.
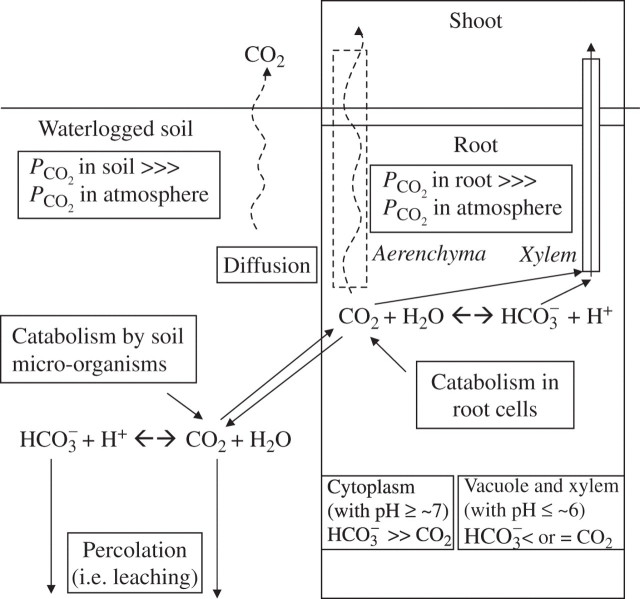
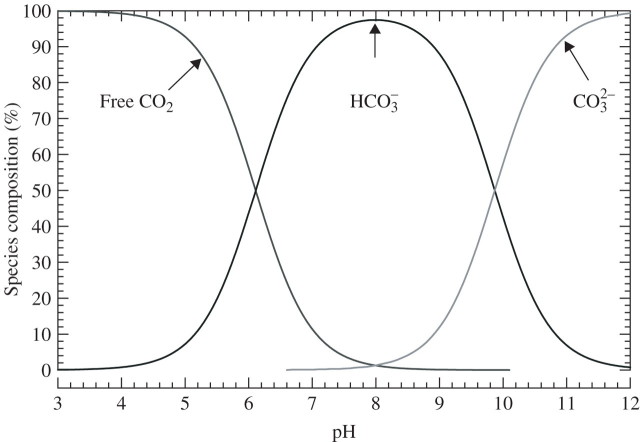
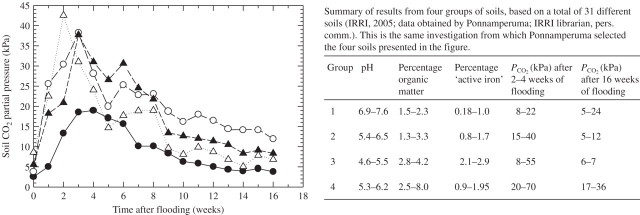
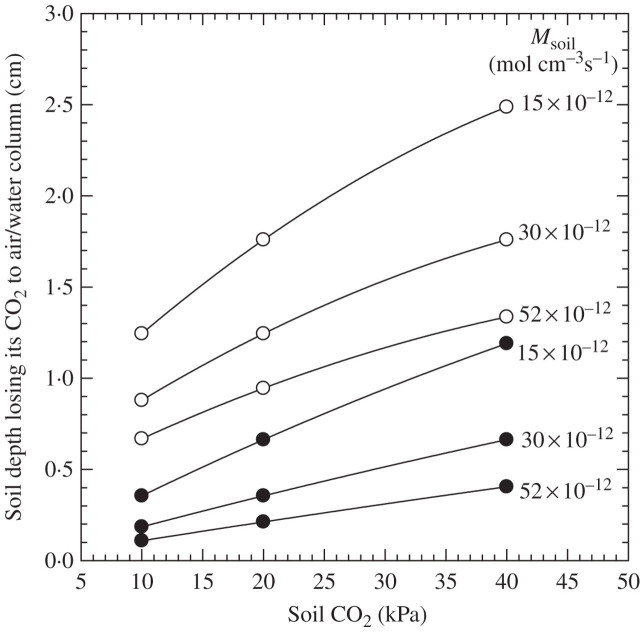
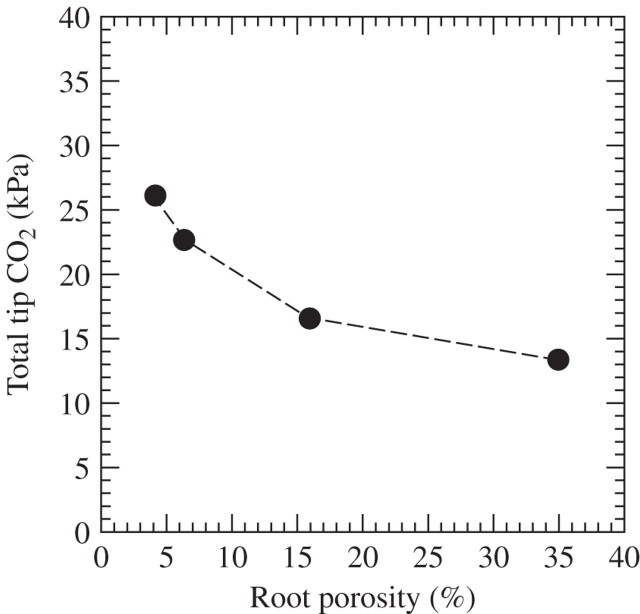
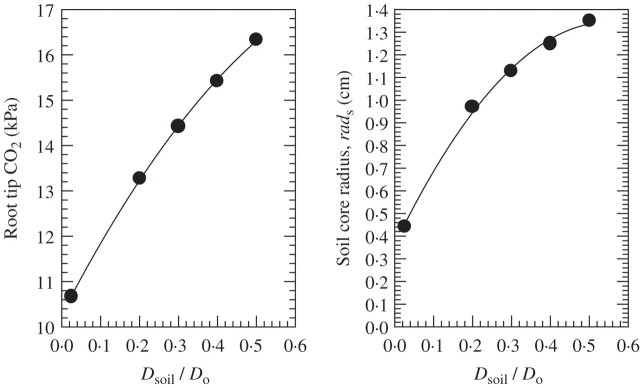
Scope: P(CO2) at 15-70 kPa has been reported for flooded paddy-field soils; however, even 15 kPa P(CO2) may not always be reached, e.g. when soil pH is above 7. Increases of P(CO2) in soils following waterlogging will develop much more slowly than decreases in P(O2); in soil from rice paddies in pots without plants, maxima in P(CO2) were reached after 2-3 weeks. There are no reliable data on P(CO2) in roots when in waterlogged or flooded soils. In rhizomes and internodes, P(CO2) sometimes reached 10 kPa, inferring even higher partial pressures in the roots, as a CO2 diffusion gradient will exist from the roots to the rhizomes and shoots. Preliminary modelling predicts that when P(CO2) is higher in a soil than in roots, P(CO2) in the roots would remain well below the P(CO2) in the soil, particularly when there is ventilation via a well-developed gas-space continuum from the roots to the atmosphere. The few available results on the effects of P(CO2) at > 5 kPa on growth have nearly all involved sudden increases to 10-100 kPa P(CO2); consequently, the results cannot be extrapolated with certainty to the much more gradual increases of P(CO2) in waterlogged soils. Nevertheless, rice in an anaerobic nutrient solution was tolerant to 50 kPa CO2 being suddenly imposed. By contrast, P(CO2) at 25 kPa retarded germination of some maize genotypes by 50%. With regard to metabolism, assuming that the usual pH of the cytoplasm of 7.5 was maintained, every increase of 10 kPa CO2 would result in an increase of 75-90 mM HCO3(-) in the cytoplasm. pH maintenance would depend on the biochemical and biophysical pH stats (i.e. regulatory systems). Furthermore, there are indications that metabolism is adversely affected when HCO3(-) in the cytoplasm rises above 50 mM, or even lower; succinic dehydrogenase and cytochrome oxidase are inhibited by HCO3(-) as low as 10 mM. Such effects could be mitigated by a decrease in the set point for the pH of the cytoplasm, thus lowering levels of HCO3(-) at the prevailing P(CO2) in the roots.
Conclusions: Measurements are needed on P(CO2) in a range of soil types and in roots of diverse species, during waterlogging and flooding. Species well adapted to high P(CO2) in the root zone, such as rice and other wetland plants, thrive even when P(CO2) is well over 10 kPa; mechanisms of adaptation, or acclimatization, by these species need exploration.
Figures
References
-
- Ahuja LR, Rawls WJ, Nielsen DR, Williams RD. 1999. Determining soil hydraulic properties and their field variability from simpler measurements. In: Skaggs RW, van Schilfgaarde J, eds. Agricultural drainage. Madison: American Society of Agronomy, 1207–1233.
-
- Alberts B, Johnson A, Lewis J, Raff M, Roberts K. 2002. Molecular biology of the cell, 4th edn. New York: Garland Science Publishers.
-
- Amthor JS. 1997. Plant respiratory responses to elevated carbon dioxide partial pressure. In: Allen LH, Kirkham MB, Olszyk DM, Whitman CE, eds. Advances in carbon dioxide effects research. Madison: American Society of Agronomy, 35–77.
-
- Armstrong W. 1979. Aeration in higher plants. Advances in Botanical Research 7: 225–332.
