

Central pathways of pulmonary and lower airway vagal afferents
- PMID: 16645192
- PMCID: PMC4503231
- DOI: 10.1152/japplphysiol.00252.2006
Central pathways of pulmonary and lower airway vagal afferents
Abstract
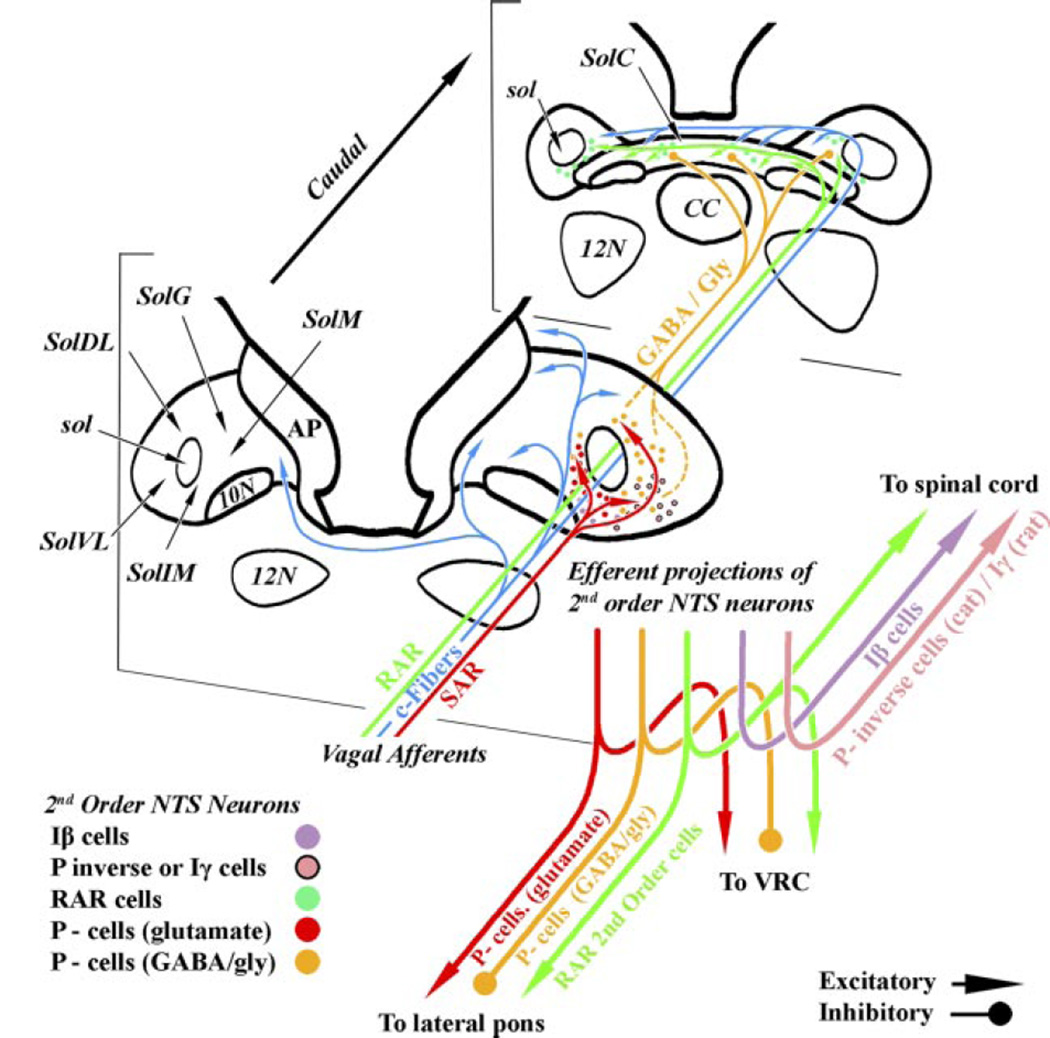
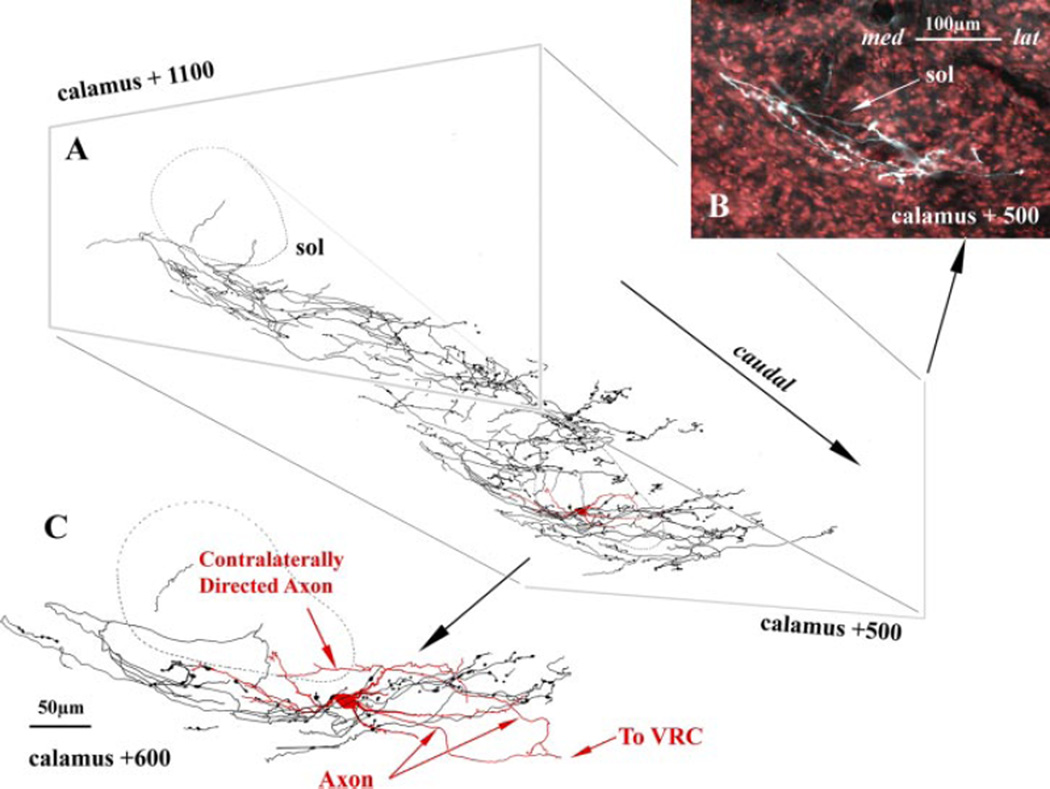
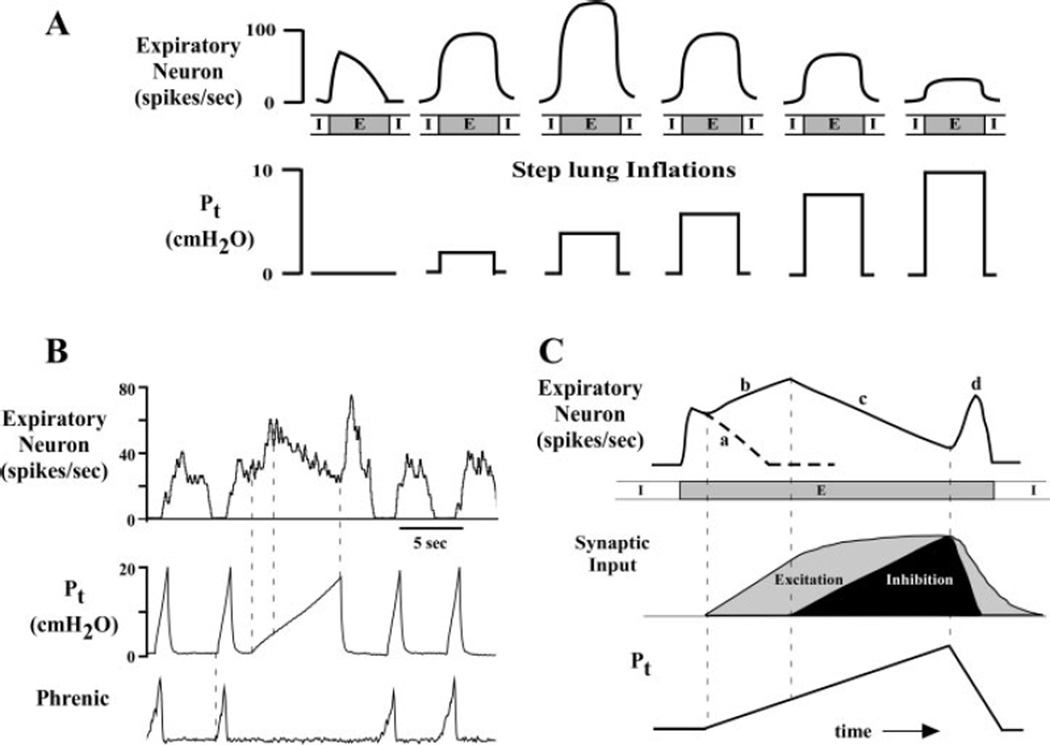
Lung sensory receptors with afferent fibers coursing in the vagus nerves are broadly divided into three groups: slowly (SAR) and rapidly (RAR) adapting stretch receptors and bronchopulmonary C fibers. Central terminations of each group are found in largely nonoverlapping regions of the caudal half of the nucleus of the solitary tract (NTS). Second order neurons in the pathways from these receptors innervate neurons located in respiratory-related regions of the medulla, pons, and spinal cord. The relative ease of selective activation of SARs, and to a lesser extent RARs, has allowed for more complete physiological and morphological characterization of the second and higher order neurons in these pathways than for C fibers. A subset of NTS neurons receiving afferent input from SARs (termed pump or P-cells) mediates the Breuer-Hering reflex and inhibits neurons receiving afferent input from RARs. P-cells and second order neurons in the RAR pathway also provide inputs to regions of the ventrolateral medulla involved in control of respiratory motor pattern, i.e., regions containing a predominance of bulbospinal premotor neurons, as well as regions containing respiratory rhythm-generating neurons. Axon collaterals from both P-cells and RAR interneurons, and likely from NTS interneurons in the C-fiber pathway, project to the parabrachial pontine region where they may contribute to plasticity in respiratory control and integration of respiratory control with other systems, including those that provide for voluntary control of breathing, sleep-wake behavior, and emotions.
Figures
References
-
- Averill DB, Cameron WE, Berger AJ. Monosynaptic excitation of dorsal medullary respiratory neurons by slowly adapting pulmonary stretch receptors. J Neurophysiol. 1984;52:771–785. - PubMed
-
- Backman SB, Anders C, Ballantyne D, Röhrig N, Camerer H, Mifflin S, Jordan D, Dickhaus H, Spyer KM, Richter DW. Evidence for a monosynaptic connection between slowly adapting pulmonary stretch receptor afferents and inspiratory beta neurones. Pflügers Arch. 1984;402:129–136. - PubMed
-
- Bajic J, Zuperku EJ, Hopp FA. Processing of pulmonary afferent input patterns by respiratory I-beta neurons. Am J Physiol Regul Integr Comp Physiol. 1989;256:R379–R393. - PubMed
-
- Bajic J, Zuperku EJ, Tonkovic-Capin M, Hopp FA. Expiratory bulbospinal neurons of dogs I. Control of discharge patterns by pulmonary stretch receptors. Am J Physiol Regul Integr Comp Physiol. 1992;262:R1075–R1086. - PubMed
-
- Bajic J, Zuperku EJ, Tonkovic-Capin M, Hopp FA. Interaction between chemoreceptor and stretch receptor inputs at medullary respiratory neurons. Am J Physiol Regul Integr Comp Physiol. 1994;266:R1951–R1961. - PubMed
Publication types
MeSH terms
Grants and funding
LinkOut - more resources
Full Text Sources
Other Literature Sources
Miscellaneous