

Maintenance of DNA methylation during the Arabidopsis life cycle is essential for parental imprinting
- PMID: 16648367
- PMCID: PMC1475502
- DOI: 10.1105/tpc.106.041178
Maintenance of DNA methylation during the Arabidopsis life cycle is essential for parental imprinting
Abstract
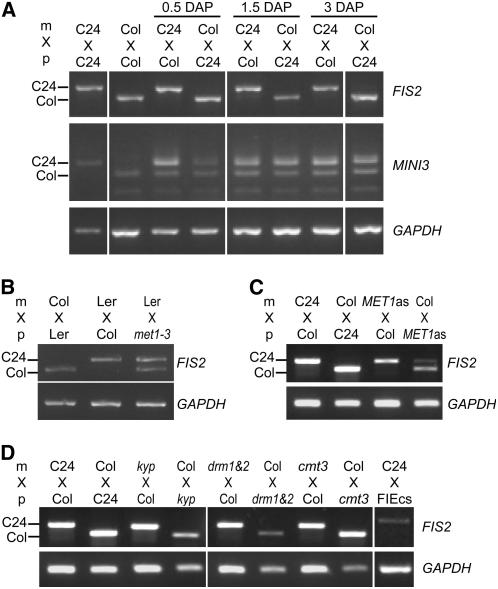
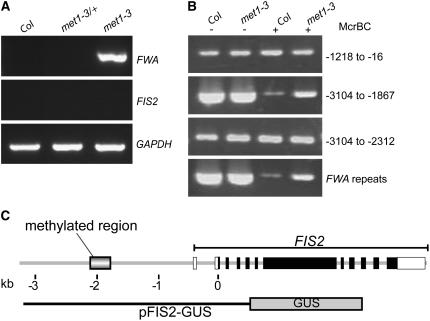
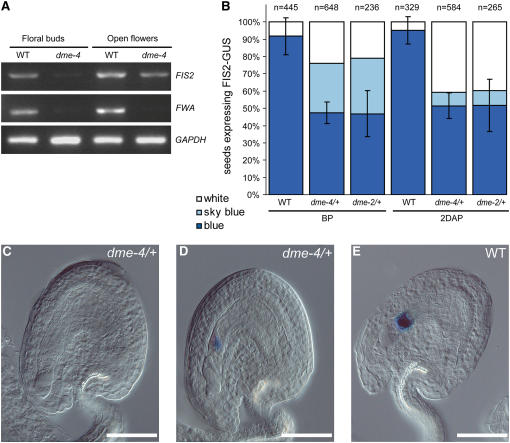
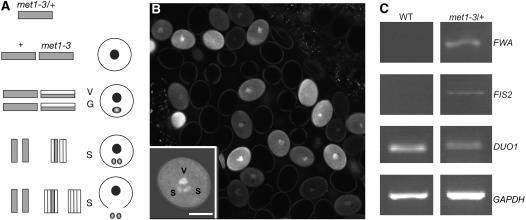
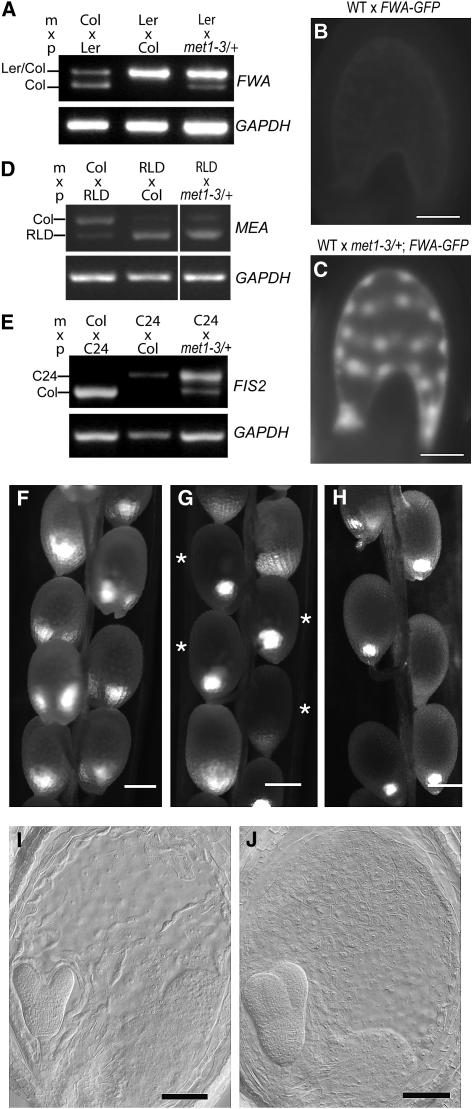
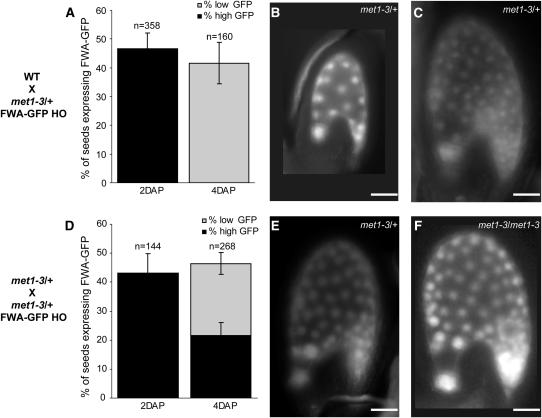
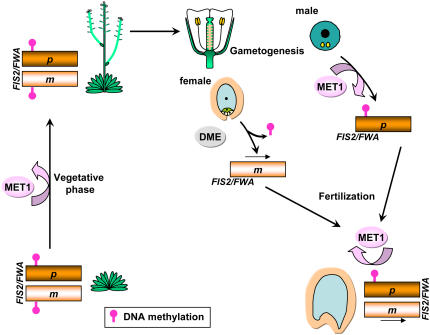
Imprinted genes are expressed predominantly from either their paternal or their maternal allele. To date, all imprinted genes identified in plants are expressed in the endosperm. In Arabidopsis thaliana, maternal imprinting has been clearly demonstrated for the Polycomb group gene MEDEA (MEA) and for FWA. Direct repeats upstream of FWA are subject to DNA methylation. However, it is still not clear to what extent similar cis-acting elements may be part of a conserved molecular mechanism controlling maternally imprinted genes. In this work, we show that the Polycomb group gene FERTILIZATION-INDEPENDENT SEED2 (FIS2) is imprinted. Maintenance of FIS2 imprinting depends on DNA methylation, whereas loss of DNA methylation does not affect MEA imprinting. DNA methylation targets a small region upstream of FIS2 distinct from the target of DNA methylation associated with FWA. We show that FWA and FIS2 imprinting requires the maintenance of DNA methylation throughout the plant life cycle, including male gametogenesis and endosperm development. Our data thus demonstrate that parental genomic imprinting in plants depends on diverse cis-elements and mechanisms dependent or independent of DNA methylation. We propose that imprinting has evolved under constraints linked to the evolution of plant reproduction and not by the selection of a specific molecular mechanism.
Figures
References
-
- Adams, S., Vinkenoog, R., Spielman, M., Dickinson, H.G., and Scott, R.J. (2000). Parent-of-origin effects on seed development in Arabidopsis thaliana require DNA methylation. Development 127 2493–2502. - PubMed
-
- Bastow, R., Mylne, J.S., Lister, C., Lippman, Z., Martienssen, R.A., and Dean, C. (2004). Vernalization requires epigenetic silencing of FLC by histone methylation. Nature 427 164–167. - PubMed
-
- Berger, F. (2003). Endosperm: The crossroad of seed development. Curr. Opin. Plant Biol. 6 42–50. - PubMed
-
- Berger, F. (2004). Plant sciences. Imprinting—A green variation. Science 303 483–485. - PubMed
-
- Cao, X., and Jacobsen, S.E. (2002. a). Role of the Arabidopsis DRM methyltransferases in de novo DNA methylation and gene silencing. Curr. Biol. 12 1138–1144. - PubMed
Publication types
MeSH terms
Substances
LinkOut - more resources
Full Text Sources
Molecular Biology Databases
