

Erasure of histone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance
- PMID: 16648464
- PMCID: PMC1472903
- DOI: 10.1101/gad.1417706
Erasure of histone acetylation by Arabidopsis HDA6 mediates large-scale gene silencing in nucleolar dominance
Abstract
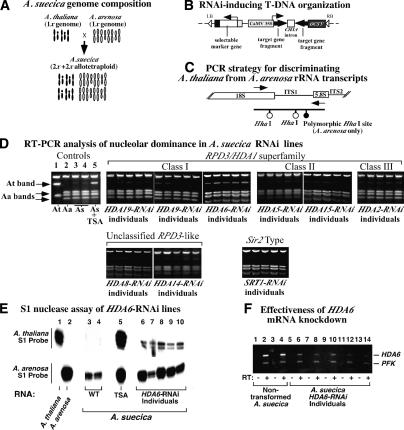
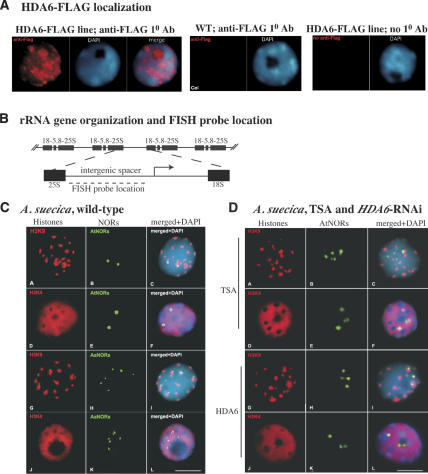
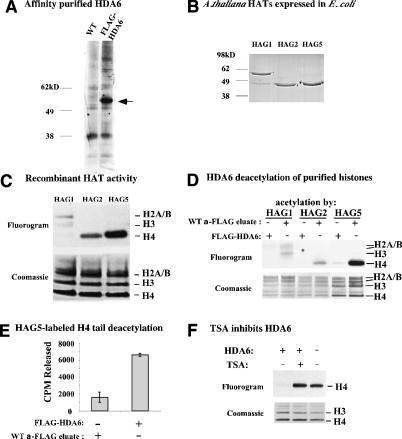
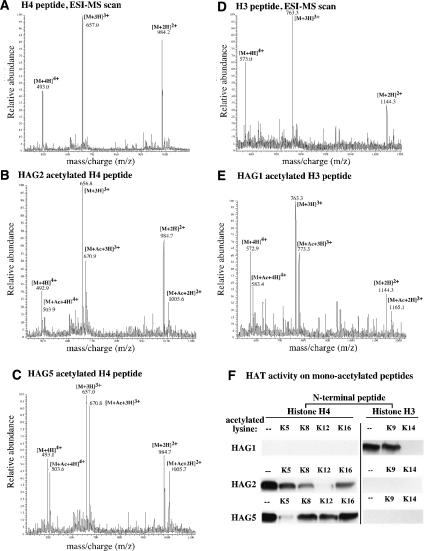
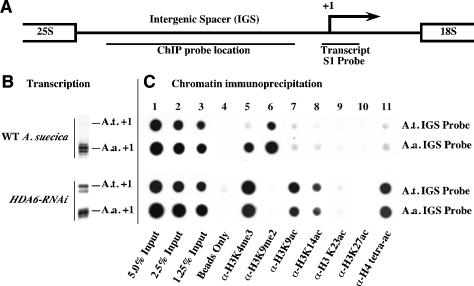
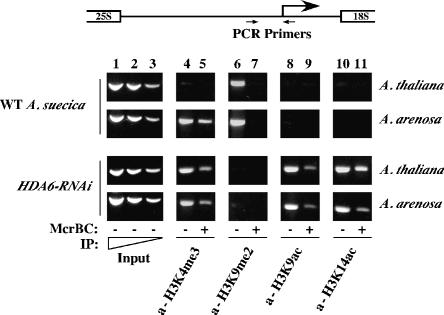
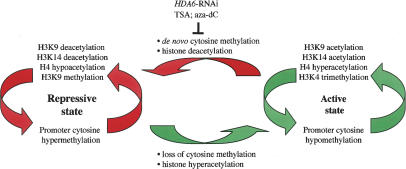
Nucleolar dominance describes the silencing of one parental set of ribosomal RNA (rRNA) genes in a genetic hybrid, an epigenetic phenomenon that occurs on a scale second only to X-chromosome inactivation in mammals. An RNA interference (RNAi) knockdown screen revealed that the predicted Arabidopsis histone deacetylase, HDA6, is required for rRNA gene silencing in nucleolar dominance. In vivo, derepression of silenced rRNA genes upon knockdown of HDA6 is accompanied by nucleolus organizer region (NOR) decondensation, loss of promoter cytosine methylation, and replacement of histone H3 Lys 9 (H3K9) dimethylation with H3K4 trimethylation, H3K9 acetylation, H3K14 acetylation, and histone H4 tetra-acetylation. Consistent with these in vivo results, purified HDA6 deacetylates lysines modified by histone acetyltransferases whose substrates include H3K14, H4K5, and H4K12. HDA6 localizes, in part, to the nucleolus, supporting a model whereby HDA6 erases histone acetylation as a key step in an epigenetic switch mechanism that silences rRNA genes through concerted histone and DNA modifications.
Figures
References
-
- Aravind L., Koonin E.V. Second family of histone deacetylases. Science. 1998;280:1167a.
-
- Berk A.J., Sharp P.A. Sizing and mapping of early adenovirus mRNAs by gel electrophoresis of S1 endonuclease-digested hybrids. Cell. 1977;12:721–732. - PubMed
-
- Buck S.W., Sandmeier J.J., Smith J.S. RNA polymerase I propagates unidirectional spreading of rDNA silent chromatin. Cell. 2002;111:1003–1014. - PubMed
Publication types
MeSH terms
Substances
Grants and funding
LinkOut - more resources
Full Text Sources
Molecular Biology Databases